Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Методы дифференциального окрашивания хромосом
|
|
В начале 70-х гг. были разработаны методы дифференциальной окраски хромосом, которые позволяли однозначно идентифицировать каждую хромосому.
Методы были основаны на способности некоторых красителей специфически связываться с конкретными участками хромосом в зависимости от их структурно-функциональной организации. Предложенные методы выявляли линейную неоднородность (сегменты) хромосом.
На практике наибольшее применение получили методы дифференциальной окраски красителем Гимза (G-окраска) и флюоресцирующим красителем акрихином или акрихинипритом (Q-окраска) (рис. 6.5, 6.6).
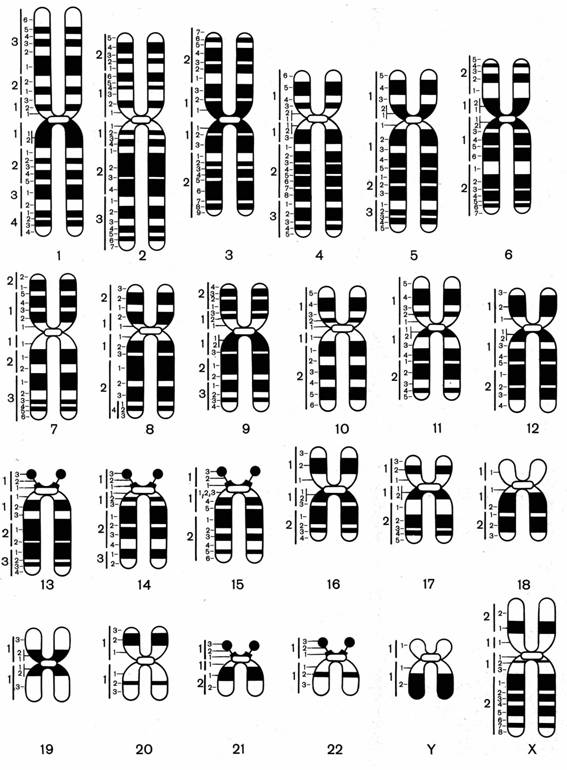
Рис. 6.5. Дифференциальная окраска хромосом
На рис. 5 представлены хромосомы человека при G-окраске. Хорошо видно, что каждая хромосома человека имеет только ей свойственную последовательность разношироких полос. Это позволяет точно идентифицировать любую из хромосом и обнаруживать относительно крупные изменения в их структуре.
При анализе метафазных хромосом средней конденсации можно четко различить около 350-400 относительно крупных сегментов на гаплоидный набор.
На стадиях, предшествующих метафазе, хромосомы менее спирализованы и поэтому имеют большую поперечную подразделенность.
Были разработаны методы анализа хромосом на делящихся клетках в стадии прометафазы. Использование этого методического подхода позволило получить хромосомы с разной степенью сегментации — от 800 до 2500 сегментов на гаплоидный набор.
На рис. 6 представлены дифференциально окрашенные X и Y-хромосомы с различными уровнями спирализации. Использованный подход дает возможность точно устанавливать точки разрывов в перестроенных хромосомах, даже если в перестройку вовлечены небольшие участки хромосом.
Поперечная исчерченность, обнаруживаемая различными методами, в принципе выявляет одни и те же сегменты хромосомы и является результатом неравномерной конденсации хроматина по всей ее длине. В зависимости от степени спирализации ДНК в хромосоме выделяют гетерохроматиновые и эухроматиновые районы, для которых характерны различные функциональные и генетические свойства.
Гетерохроматиновый район представляет собой участок конденсированного хроматина (высокоспирализованная ДНК), который выявляется при дифференциальном окрашивании в виде темных полос. Присутствие гетерохроматина можно обнаружить и в интерфазном ядре, где он отчетливо выявляется в виде интенсивно окрашенных глыбок хроматина. Считывания генетической информации с данных участков не происходит.
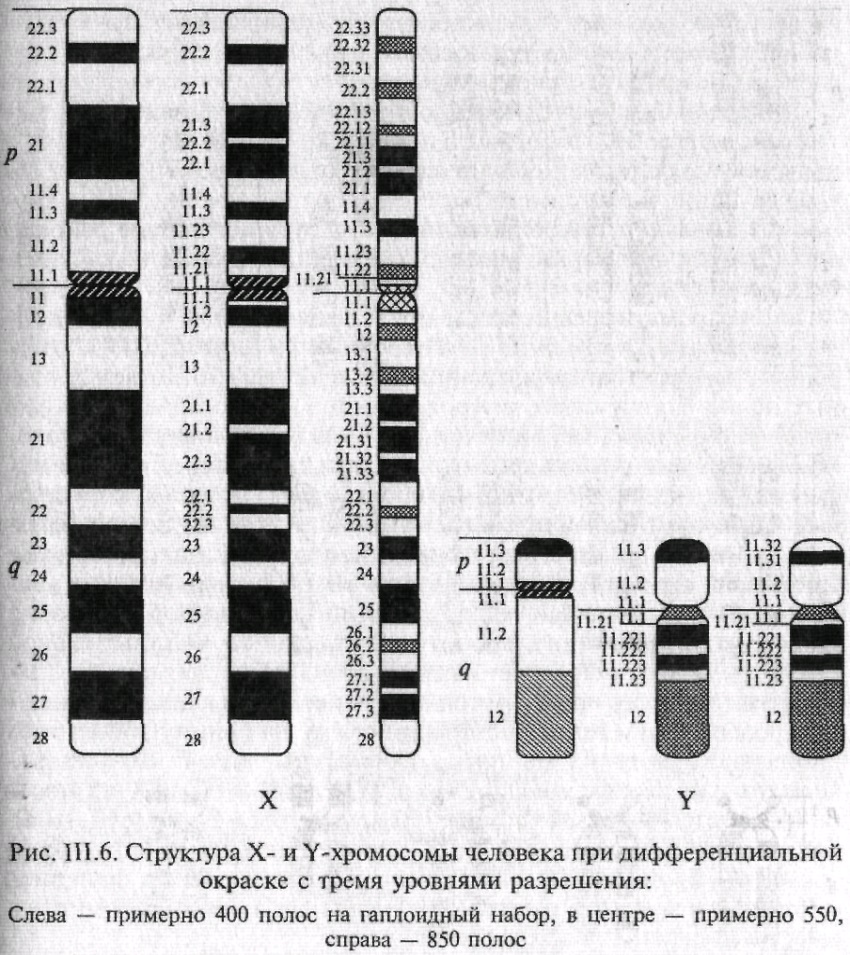
Рис. 6.6. Структура хромосом человека при дифференциальной окраске с тремя уровнями разрешения
Различают структурный и факультативный гетерохроматин. Структурный гетерохроматин постоянно присутствует в определенных регионах хромосомы. Например, он всегда обнаруживается вокруг центромер всех хромосом. Факультативный гетерохроматин появляется в хромосоме при сверхспирализации эухроматиновых районов. Факультативной гетерохроматизацией может быть охвачена целая хромосома.
Так, в клетках женского организма одна из Х-хромосом полностью инактивирована путем гетерохроматизации уже на ранних этапах эмбрионального развития. Ее можно обнаружить в виде глыбки гетерохроматина на периферии ядра. Такая инактивированная Х-хромосома называется половым хроматином, или тельцем Барра.
Благодаря гетерохроматизации Х-хромосомы в клетках женского организма происходит выравнивание количества генов, функционирующих в мужском и женском организмах, поскольку у мужчин имеется только одна Х-хромосома.
Эухроматиновые регионы хромосом в интерфазном ядре не видны, поскольку представлены хроматином в деконденсированном состоянии. Это указывает на их высокую метаболическую активность. Действительно, эухроматиновые районы содержат уникальные гены, контролирующие синтез различных белков. При дифференциальном окрашивании метафазных хромосом они определяются как светлые полосы.
Изучение химического состава хромосом показало, что они состоят из дезоксирибонуклеиновой кислоты (ДНК), гистонов, негистонных белков и небольшого количества РНК.
В каждой хромосоме содержится только одна молекула ДНК. Однако размеры молекул ДНК хромосом огромны. Они могут достигать сотен микрометров и даже сантиметров. В кариотипе человека самая большая хромосома — 1; длина ее полностью раскрученной ДНК составляет около 7 см, что значительно больше не только размеров ядра клетки, но и самой клетки. Суммарная длина молекул ДНК всех хромосом одной клетки человека составляет более 170 см. Несмотря на свои гигантские размеры, молекулы ДНК функционируют в пределах таких микрообразований, как хромосомы. Поэтому хромосомы ядер клеток должны представлять собой сильно укороченные (конденсированные) структуры ДНК. Это достигается за счет специфической укладки молекул ДНК — многоуровневой спирализации. Основной структурной единицей хромосомы является нуклеосома (рис. 6.7, 6.8).
Каждая нуклеосома содержит по две молекулы четырех различных типов гистонов, объединенных в октамер (восьмигранник), обвитый нитью ДНК. Нуклеосомы и соединяющие их участки ДНК формируют спиральную структуру — хроматиновое волокно. На каждый виток такой спирали приходится 6 нуклеосом. Так формируется структура хромосомы (рис. 6.7, 6.8).
При конденсации происходит уменьшение длины молекулы ДНК в 10 тыс. раз, так что конденсированные хромосомы в среднем имеют длину порядка 200 нм (т. е. 200х10-9 м). Это обеспечивает возможность точного и быстрого деления генетического материала материнской клетки между дочерними клетками (митоз) и уменьшение числа хромосом вдвое при образовании половых клеток (мейоз).
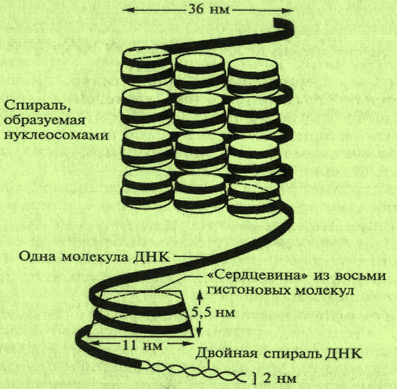
Рис. 6.7. Структура нуклеосомы
Хромосомы выполняют функцию основного генетического аппарата клетки. В них в линейном порядке расположены гены из которых занимает строго определенное место, называемое локусом. Альтернативные формы гена (т.е. различные его состояния), занимающие один и тот же локус, называются аллелями (от греч. allelon — взаимно другой, иной). Любая хромосома содержит только единственный аллель в данном локусе, несмотря на то, что в популяции могут существовать два, три и более аллелей одного гена.
Уровни организации хроматина (рис. 6.8)
• метафазная хромосома – 1400 нм
• участок метафазной хромосомы – 700 нм;
нуклеомеры- субъединицы толстой фибриллы - 300 нм
• хроматиновая фибрилла – 30 нм;
• нуклеосомы – 11 нм
• двойная спираль ДНК – 2 нм
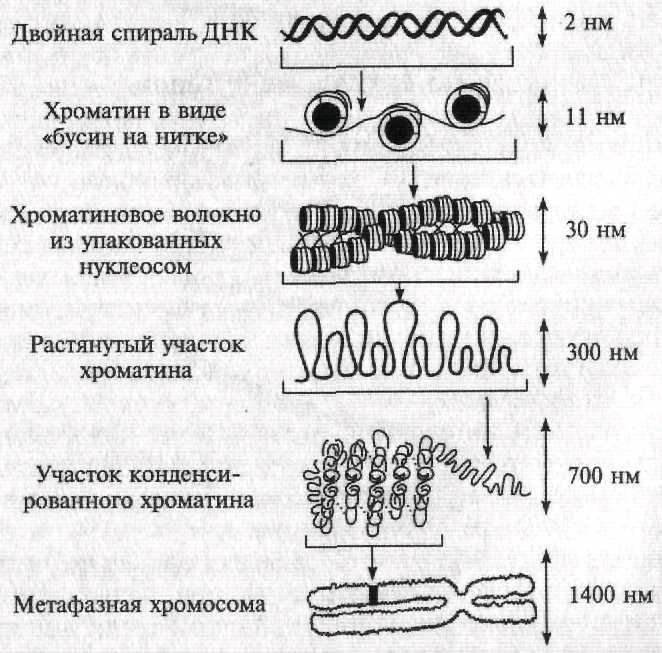
Рис. 6.8. Уровни организации хроматина