Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Модуль 1. 1)Потенціал спокою (ПС) – різниця потенціалів між зовнішньою та внутрішньою поверхнями мембрани збудливої клітини
|
|
1) Потенціал спокою (ПС) – різниця потенціалів між зовнішньою та внутрішньою поверхнями мембрани збудливої клітини, що знаходиться в стані спокою.
Формула Нернста для розрахунку величини ПС: 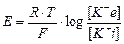 , де
, де
R – універсальна газова стала, T – абсолютна температура, F – число Фарадея, [K+ і] – концентрація іонів К+ в клітині, [K+ е] – концентрація іонів К+ поза клітиною.
Механізм формування ПС пов’язаний з:
1. Наявністю в мембрані клітини механізмів активного транспорту речовин – натрій-калієвий насос. Він створює градієнт концентрації усередині та поза клітиною.
2. Особливостями проникності мембрани клітини в стані спокою – вона проникна для іонів К+ та непроникна для іонів Na+.
 Найбільш важливим для створення та підтримання ПС є натрій-калієвий насос (НКН). Структурною одицею НКН є Na/К-транспортний білок, що являє собою АТФазу. Цей білок на внутрішній поверхні мембрани розщеплює АТФ на АДФ та фосфат й використовує енергію, що виділилась на транспортування трьох іонів Na+ з клітини та двох іонів К+ в клітину. Тобто в сумі з клітини за один цикл виділяється один позитивний заряд (150-600 за секунду). Таким чином, НКН є електрогенним (створює електричний струм через мембрану), але й може бути електронейтральним, коли на один іон транспортованого Na+ припадає один іон К+. Отже, за рахунок роботи НКН створюється та підтримується градієнт концентрації іонів К+ та іонів Na+ усередині та поза клітиною, а саме 145 ммоль/л іонів Na+ зовні та 12 ммоль/л всередині, 4 ммоль/л іонів К+ зовні та 155 ммоль/л всередині. Отже, концентрація іонів калію усередині клітини в 40-50 разів більша, чим поза нею, а концентрація іонів натрію, навпаки, в 10 разів менша. Варто підкреслити, що сумарна концентрація аніонів усередині та поза клітиною однакова.
Найбільш важливим для створення та підтримання ПС є натрій-калієвий насос (НКН). Структурною одицею НКН є Na/К-транспортний білок, що являє собою АТФазу. Цей білок на внутрішній поверхні мембрани розщеплює АТФ на АДФ та фосфат й використовує енергію, що виділилась на транспортування трьох іонів Na+ з клітини та двох іонів К+ в клітину. Тобто в сумі з клітини за один цикл виділяється один позитивний заряд (150-600 за секунду). Таким чином, НКН є електрогенним (створює електричний струм через мембрану), але й може бути електронейтральним, коли на один іон транспортованого Na+ припадає один іон К+. Отже, за рахунок роботи НКН створюється та підтримується градієнт концентрації іонів К+ та іонів Na+ усередині та поза клітиною, а саме 145 ммоль/л іонів Na+ зовні та 12 ммоль/л всередині, 4 ммоль/л іонів К+ зовні та 155 ммоль/л всередині. Отже, концентрація іонів калію усередині клітини в 40-50 разів більша, чим поза нею, а концентрація іонів натрію, навпаки, в 10 разів менша. Варто підкреслити, що сумарна концентрація аніонів усередині та поза клітиною однакова.
Проникність мембрани для іонів визначається станом каналів мембрани. Стан каналів визначається станом їх воріт, які можуть бути відкритими (тоді через канал по градієнту концентрації можуть рухатися іони) та закритими. Канали мембрани є селективними, тобто, пропускають одні іони та не пропускають, або дуже погано пропускають інші іони. В стані спокою мембрана збудливої клітини проникна для іонів К+ (відкриті калієві канали) та непроникна для іонів Na+ – майже всі натрієві канали закриті. Це є причиною того, що іони К+ виходять з клітини через калієві канали по градієнту концентрації, а аніони Na+ зайти в клітину не можуть, хоча є градієнт концентрації. Іони К+, виходячи з клітини, виносять позитивний заряд на зовнішню поверхню мембрани. Крупномолекулярні органічні аніони залишаються в клітині й зумовлюють негативний заряд на внутрішній поверхні мембрани.
 |
|
|
При реєстрації ПС методом пошкодження об’єктом дослідження є який-небудь орган, наприклад, скелетний м’яз. Його пошкоджують шляхом розрізу та прикладають електроди до пошкодженої та непошкодженої поверхонь. Стрілка гальванометра при цьому відхиляється. Це говорить про наявність різниці потенціалів між пошкодженою поверхнею м’яза (аналог внутрішньоклітинного середовища) та її непошкодженою поверхонею (аналог зовнішньої мембрани).
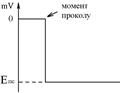 При внутрішньоклітинному методі реєстрації об’єктом дослідження є окрема клітина, яку поміщають в плазму чи фізіологічний розчин. Там знаходиться один реєструючий електрод. Другий електрод (мікроелектрод) обережно вводиться усередину клітини. До проколу мембрани промінь осцилографа іде по нульовій лінії Ù між електродами немає різниці потенціалів. В момент проколу мембрани промінь приладу різко відхиляється від 0 у негативну частину системи координат і протягом годин може перебувати на такому рівні.
При внутрішньоклітинному методі реєстрації об’єктом дослідження є окрема клітина, яку поміщають в плазму чи фізіологічний розчин. Там знаходиться один реєструючий електрод. Другий електрод (мікроелектрод) обережно вводиться усередину клітини. До проколу мембрани промінь осцилографа іде по нульовій лінії Ù між електродами немає різниці потенціалів. В момент проколу мембрани промінь приладу різко відхиляється від 0 у негативну частину системи координат і протягом годин може перебувати на такому рівні.
Внутрішньоклітинна реєстрація ПС дає змогу оцінити його параметри:
1. Полярність – те, що в момент проколу промінь відхиляється у негативну сторону, говорить про те, щовнутрішня поверхня мембрани клітини заряджена негативно відносно зовнішньої.
2. Величина – ступінь відхилення променя від нуля, відповідає величині ПС. Величина ПС збудливих клітин складає 70-90 мВ. Варто наголосити, що величина ПС оцінюється по модулю, без врахування знака!
3. Сталість у часі – якщо, на клітину не діють які-небудь фактори, то ПС не змінюється (промінь йде паралельно нульовій лінії).
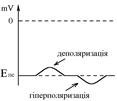 Так як величина ПС оцінюється по модулю, то деполяризацією називають будь-яке зменшення величини ПС, а гіперполяризацією – його збільшення.
Так як величина ПС оцінюється по модулю, то деполяризацією називають будь-яке зменшення величини ПС, а гіперполяризацією – його збільшення.
Фізіологічна роль ПС полягає в тому, що він є базою, на якій розвивається потенціал дії.
2 ) Потенціал дії (ПД) – швидка високоамплітудна зміна потенціалу мембрани збудливої клітини при її збуджені.
| |||||||||||||||||
 | |||||||||||||||||
 | |||||||||||||||||
|
| ||||||||||||||||
 |  | ||||||||||||||||
|
|
При позаклітинній реєстрації об’єктом досліджень є цілий нерв чи м’яз. При біполярному відведені реєструється двохфазний сумарний ПД (обидва електроди є активними, дві фази ПД відповідають руху хвилі збудження спочатку під одним, а потім – під іншим електродом). При уніполярній реєстрації один електрод є активним, а інший – пасивним Ù реєструється однофазний сумарний ПД. Вихідною лінією при позаклітинній реєстрації є нульова лінія, що свідчить про відсутність різниці потенціалів між електродами.
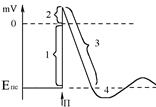 При внутрішньоклітинній (мікроелектродній) реєстрації ПД окремої клітини має такий вигляд:
При внутрішньоклітинній (мікроелектродній) реєстрації ПД окремої клітини має такий вигляд:
Спочатку мембранний потенціал різко зменшується до нуля – 1 фаза деполяризації ПД; потім заряд мембрани змінюється на протилежний – зовні “-”, всередині “+” – 2 фаза реверсполяризації. Частина графіка, яка перебуває вище нуля відповідає фазі, що має назву овершут. Далі мембранний потенціал поступово повертається до вихідного рівня – 3 фаза реполяризації ПД. Завершальною фазою ПД є слідова гіперполяризація 4.
В основі зміни мембранного потенціалу при збуджені (тобто в основі формуванні ПД) лежать зміни проникності мембрани, що пов’язані із змінами стану каналів мембрани під впливом подразника.
 Особливістю натрієвих каналів є наявність у них двох воріт:
Особливістю натрієвих каналів є наявність у них двох воріт:
- активаційних – закриті в стані спокою, але можуть відкриватися під впливом збудника;
- інактиваційних – вони швидко закриваються після відкриття активаційних воріт і не можуть відкриватися під впливом подразника.
Подразник зумовлює відкриття активаційних воріт натрієвих каналів Ù лавиноподібний вхід Na+ в клітину за градієнтом концентрації Ù зменшення надлишку аніонів усередині клітини (так як іони Na+ заряджені позитивно). Аніони, які прямують до мембрани за іонами Na+, зайти в клітину не можуть Ù вони зменшують надлишок позитивних зарядів на зовнішній поверхні мембрани клітини. Завдяки цьому різниця потенціалів між зовнішньою та внутрішньою поверхнями мембрани зменшується, а далі й зовсім зникає. Так розвивається фаза деполяризації ПД.
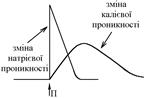 Але вхід іонів Na+ в клітину не припиняється й тепер у клітині створюється надлишок позитивних іонів Na+, а на її поверхні – надлишок аніонів Ù перезарядка мембрани – фаза реверсполяризації.
Але вхід іонів Na+ в клітину не припиняється й тепер у клітині створюється надлишок позитивних іонів Na+, а на її поверхні – надлишок аніонів Ù перезарядка мембрани – фаза реверсполяризації.
Натрієва активація (стан каналів при відкритих активаційних воротах) швидко змінюється натрієвою інактивацією – закриттям інактиваційних воріт Ù вхід іонів Na+ в клітину при цьому припиняється. Подразник змінює не тільки стан натрієвих каналів, але й стан калієвих каналів Ù вони також активуються, тобто відкривається додаткова кількість калієвих каналів. Але у часі цей процес розвивається значно повільніше, ніж натрієва активація.
Внаслідок різної швидкості розвитку у часі натрієвої та калієвої активації під впливом подразника, розвиток натрієвої інактивації супроводжується розвитком калієвої активації. Тому, коли іони Na+ перестають входити в клітину, вихід іонів К+ з неї збільшується. Саме це призводить до відновлення вихідного рівня мембранного потенціалу – фаза реполяризації.
Таким чином механізм формування ПД заключається в наступному:
 | |||||||||||||||||||
 | |||||||||||||||||||
 | |||||||||||||||||||
 |  | | |||||||||||||||||
|  | ||||||||||||||||||
| | | |||||||||||||||||
|
|
| |||||||||||||||||
Параметрами ПД є:
1.Амплітуда – 100-120 мВ.
2.Протяжність – 1-3 мс в нервових волокнах, в м’язових волокнах він довший (в типових кардіоміоцитах – 300 мс).
Фізіологічна роль ПД – полягає в тому, що він забезпечує збудження клітини та передачу цього збудження на інші клітини, а також підтримку чи активацію у збудливих клітинах специфічних функцій.
3) Механізми проведення збудження по нервових волокнам
За будовою всі нервові волокна поділяють на:
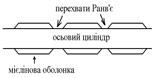 - безмієлінові – мієлінової оболонки не мають;
- безмієлінові – мієлінової оболонки не мають;
- мієлінові – мають мієлінову оболонку; при цьому певні частини волокна вкриті мієліновою оболонкою, а між ними є невкриті проміжки – перехвати Ранв’є; особливістю мієлінової оболонки є її високий опір – вона складається з фосфоліпідів, які є диелектриками (ізоляторами).
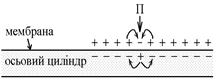 Механізми проведення збудження по безмієліновим нервовим волокнам такий. Під впливом подразника (П) на мембрані волокна виникає збудження – ПД (на рисунку зовні “-”, всередині “+” – показана фаза реверсполяризації ПД – максимум розвитку піка). Між збудженими та незбудженими (зовні “+”, всередині “-” – потенціал спокою) ділянками мембрани волокна виникає різниця потенціалів, Δ U; зовні та всередині ці ділянки сполучені провідним середовищом (цитоплазма та міжклітинна рідина) Ù між цими ділянками мембрани виникають місцеві елекричні струми, що спрямовані від “+” до “-“, діють на мембрану із зовнішньої та внутрішньої поверхні. Ці струми є подразниками для мембрани. Необхідно оцінити параметри цих струмів як подразника:
Механізми проведення збудження по безмієліновим нервовим волокнам такий. Під впливом подразника (П) на мембрані волокна виникає збудження – ПД (на рисунку зовні “-”, всередині “+” – показана фаза реверсполяризації ПД – максимум розвитку піка). Між збудженими та незбудженими (зовні “+”, всередині “-” – потенціал спокою) ділянками мембрани волокна виникає різниця потенціалів, Δ U; зовні та всередині ці ділянки сполучені провідним середовищом (цитоплазма та міжклітинна рідина) Ù між цими ділянками мембрани виникають місцеві елекричні струми, що спрямовані від “+” до “-“, діють на мембрану із зовнішньої та внутрішньої поверхні. Ці струми є подразниками для мембрани. Необхідно оцінити параметри цих струмів як подразника:
1. Напрям – з рисунка видно, що струми мають вихідний (катодний) напрям в незбуджених ділянках мембрани. Отже, тут буде виникати деполяризація мембрани. Якщо вона дійде до Екр, то виникне ПД.
2. Сила – чисельно сила струму в даному випадку рівна різниці потенціалів між збудженими та незбудженими ділянками мембрани і ця сила відповідає амплітуді ПД. Поріг сили відповідає величині порогового потенціалу (Δ V). Амплітуда ПД нервового волокна складає 100-120 мВ, поріг деполяризації – 15-20 мВ. Відповідно, сила подразника в декілька разів (6-8) більша порогової. Відношення амплітуди ПД до порогу деполяризації має назву коефіцієнт надійності і показує, у скільки разів сила місцевого струму як подразника більша за порогову силу.
3. Час дії подразника – відповідає тривалості ПД і в декілька разів більший порогового.
4. Швидкість збільшення сили – відповідає швидкості піку ПД, майже, відповідає швидкості збільшення сили при дії прямокутних імпульсів електричного струму – набагато вища порогу.
Отже, на збуджену ділянку мембрани нервового волокна діє катодний електричний струм, сила, час дії та швидкість збільшення сили, якого вищі порогу – цей струм викличе деполяризацію мембрани до ЕкрÙ викличе ПД на мембрані незбудженої ділянки.
Далі процес повторюється Ù ПД поширюється вздовж всієї мембрани нервового волокна.
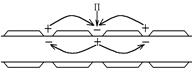 Механізм проведення збудження по мієліновим нервовим волокнам принципово не відрізняється від механізму проведення по безмієлінових волокнах: під впливом подразника (П) в одному з перехватів Ранв’є виникає ПД – на мембрані перезарядка Ù між цим (збудженим) і сусідніми (незбудженими) перехватами Ранв’є виникає різниця потенціалів Δ U; вони з’єднані провідним середовищем Ù виникають місцеві струми (від “+” до “-”). Ці струми в ділянці незбуджених перехватів мають вихідний напрямок; їх сила (амплітуда ПД), тривалість (тривалість ПД), швидкість збільшення сили (швидкість збільшення піку ПД) надпорогові Ù на мембрані незбудженого перехвату Ранв’є виникає деполяризація, яка досягає ЕкрÙ виникає ПД.
Механізм проведення збудження по мієліновим нервовим волокнам принципово не відрізняється від механізму проведення по безмієлінових волокнах: під впливом подразника (П) в одному з перехватів Ранв’є виникає ПД – на мембрані перезарядка Ù між цим (збудженим) і сусідніми (незбудженими) перехватами Ранв’є виникає різниця потенціалів Δ U; вони з’єднані провідним середовищем Ù виникають місцеві струми (від “+” до “-”). Ці струми в ділянці незбуджених перехватів мають вихідний напрямок; їх сила (амплітуда ПД), тривалість (тривалість ПД), швидкість збільшення сили (швидкість збільшення піку ПД) надпорогові Ù на мембрані незбудженого перехвату Ранв’є виникає деполяризація, яка досягає ЕкрÙ виникає ПД.
Суттєва відмінність поширення ПД по мієліновим волокнам полягає в тому, що в них місцеві струми виникають не між сусідніми ділянками мембрани, а між сусідніми перехватами Ранв’є Ù ПД поширюється від одного перехвату до іншого (стрибкоподібно) Ù швидкість поширення ПД збільшується.
На швидкість поширення збудження по нервовим волокнам впливають такі фактори:
1. Наявність мієлінової оболонки збільшують швидкість.
2. Відстань між перехватами Ранв’є – чим він більший, тим більша швидкість.
Так як в м’язових волокнах відсутня мієлінова оболонка, то швидкість проведення збудження по цим волокнам будуть визначати фактори 3-6.
3. Діаметр волокна – чим він більший, тим менший опір чинить аксоплазма волокна поширенню локальних струмів і тим більша швидкість проведення збудження.
4. Амплітуда ПД – чим вона більша, тим швидше деполяризація доходить до Екр, і тим більша швидкість проведення.
5. Поріг деполяризації (Δ V) – чим він менший, тим швидше деполяризація мембрани волокна доходить до Екр, і тим більша швидкість проведення.
6. Швидкість наростання піку ПД – чим вона більша, тим швидше розвивається деполяризація до Екр, і тим більша швидкість проведення збудження.
4) Закономірності проведення збудження нервовими волокнами
1. Закон фізіологічної неперервності чи фізіологічної цілісності волокна – для здійснення проведення необхідним є нормальний функціональний стан мембрани волокна. Якщо його пошкодити, обробивши, наприклад, місцевим анастетиком типу новокаїну, проведення припиниться, незважаючи на те, що морфологічно волокна не пошкоджені (місцеві анастетики інактивують натрієві канали мембрани Ù зміщення ЕкрÙ збільшення порогу деполяризації Ù зменшення швидкості проведення збудження з подальшим припиненням цього проведення).
2. Закон двостороннього проведення – в умовах експерименту збудження (ПД) поширюється вздовж волокна в дві сторони, тобто, провідність волокон двостороння. Але в умовах цілісного організму відбувається одностороннє проведення збудження по нервовим волокнам, незважаючи на двосторонню провідність – ПД поширюється від рецепторів до нервових центрів, від нервових центрів до органів-ефекторів. Це відбувається завдяки тому, що обов’язковим елементом будь-якої рефлекторної дуги є синапси, які мають односторонню провідність.
3. Закон ізольованого проведення – ПД, поширюючись по мембрані одного волокна, не передається на сусідні. Причина цього полягає в тому, що мембрана волокна має набагато більший опір, ніж міжклітинна рідина Ù місцеві струми шунтуються через міжклітинну рідину й не викликають ПД на мембрані сусідніх волокон.
4. Закон бездекрементного проведення – амплітуда ПД не змінюється при проведенні збудження по мембрані волокна. Причиною цього є те, що ПД підкоряється закону “все або нічого” – в кожній ділянці мембрани він має максимально можливу для цього волокна амплітуду.
5. Проведення без розвитку втоми. Нервові волокна практично не стомлюються – на протязі багатьох годин відтворюють частоту стимуляції. Причина цього полягає у високій економічності механізму проведення збудження по мембрані волокна – при цьому енерговитрати незначні Ù дуже низька швидкість розвитку втоми волокон.
6. Проведення з дуже високим коефіцієнтом надійності – відношення амплітуди ПД до порогу деполяризації (Δ V) в нервових волокнах дорівнює 6-8, що свідчить про велику надійність проведення збудження по цим волокнам.
7. Проведення з великою швидкістю – в товстих мієлінових нервових волокнах швидкість проведення збудження досягає 120 м за секунду.
За швидкістю проведення збудження нервові волокна поділяють на такі типи:
| Тип волокна | Діаметр волокна, мкм | Швидкість проведення, м/с |
| Аa | 100 (70-120) | |
| Аb | 50 (30-70) | |
| Аg | 20 (15-30) | |
| Аd | 15 (12-30) | |
| В | 7 (3-15) | |
| С | 1 (0, 5-2, 0) |
Волокна типу А та В – мієлінові.
Аa – первинні аференти м’язових веретен, рухові волокна скелетних м’язів.
Аb – шкірні аференти дотику та тиску.
Аg – рухові волокна м’язових веретен.
Аd – шкірні аференти температури та болю.
В – симпатичні прегангліонарні волокна.
С – безмієлінові симпатичні постгангліонарні волокна та шкірні аференти болю.
Інформація по нервовим волокнам передається точно, без змін, швидко. Ці особливості передачі інформації пов’язані з певними особливостями поширення збудження.
 | |||||||||
 | |||||||||
|
|
|
5) Механізми проведення збудження через нервово-м’язовий синапс. Медіатор, мембранні циторецептори та блокатори нервово-м’язових синапсів.
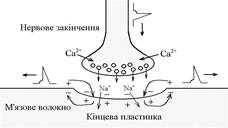 Нервово-м’язовий синапс утворений нервовим закінченням аксона a-мотонейронів та кінцевою пластинкою – частина мембрани м’язового волокна, яка контактує з нервовим закінченням. Розділяє пресинаптичну мембрану нервового волкна та кінцеву пластинку синаптична щілина.
Нервово-м’язовий синапс утворений нервовим закінченням аксона a-мотонейронів та кінцевою пластинкою – частина мембрани м’язового волокна, яка контактує з нервовим закінченням. Розділяє пресинаптичну мембрану нервового волкна та кінцеву пластинку синаптична щілина.
Механізм передачі збудження через нервово-м’язовий синапс полягає в тому, що ПД іде по мембрані нервового волокна Ù поширюється по пресинаптичній мембрані, при цьому відкриваються кальцієві канали пресинаптичної мембрани Ù вхід іонів Са+ всередину нервового закінчення Ù взаємодія з везикулами (міхурці, в яких є медіатор ацетилхолін) Ù рух везикул до пресинаптичної мембрани Ù злиття везикул з пресинаптичною мембраною Ù вихід медіатора ацетилхоліну в синаптичну щілину Ù дифузія ацетилхоліну до мембрани кінцевої пластинки Ù взаємодія з мембранними циторецепторами (Н-холінорецептори) Ù відкриття хемочутливих натрієвих каналів Ù вхід іонів Na+ в м’язове волокно через кінцеву пластинку Ù розвиток деполяризації кінцевої пластинки, що має назву потенціалу кінцевої пластинки (ПКП). ПКП – один із видів місцевого збудження. 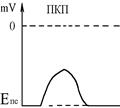
 Як будь-яке з місцевих збуджень ПКП поширюється на сусідні ділянки постсинаптичної мембрани за допомогою місцевих струмів. Ці струми в незбуджених ділянках мембрани мають вихідний напрям, тобто викликають деполяризацію.
Як будь-яке з місцевих збуджень ПКП поширюється на сусідні ділянки постсинаптичної мембрани за допомогою місцевих струмів. Ці струми в незбуджених ділянках мембрани мають вихідний напрям, тобто викликають деполяризацію.
Cила цих струмів чисельно відповідає амплітуді ПКП, а вона складає 45-50 мВ. Поріг деполяризації постсинаптичної мембрани м’язового волокна складає 40 мВ (величина ПС = –90 мВ, величина Екр = –50 мВ. Отже, сила цих струмів надпорогова.
Час дії цих струмів дорівнює тривалості ПКП і також є надпороговим.
Швидкість збільшення сили відповідає швидкості розвитку ПКП й також
є надпороговою.
Отже, місцеві струми в незбуждених ділянках постсинаптичної мембрани викликають деполяризацію до Екр Ù розвиток ПД, які будуть поширюватися від місця свого виникнення вздовж усієї довжини мембрани м’язового волокна (і через електромеханічне спряження будуть викликати скорочення м’яза).
Закономірності проведення збудження через нервово-м’язовий синапс:
1. Однобічне проведення.
2. Уповільнене проведення (час від виникнення ПД на пресинаптичній мембрані до виникнення ПД на постсинаптичній мембрані складає близько 0, 5 мсек.
3. Проведення збудження через синапс супроводжується швидким розвитком втоми (у зв’язку з виснаженням запасів медіатора в нервовому закінченні);
Ці особливості (1-3) характерні для будь-якого хімічного синапса і пов’язані з хімічним характером передачі інформації через синапс.
4. Збудження передається через нервово-м’язовий синапс без трансформації ритму: 1 ПД на пресипаптичній мембрані викликає 1 ПКП на мембрані кінцевої пластинки, а це викликає 1 ПД на мембрані м’язового волокна.
Блокаторами нервово-м’язової передачі є курареподібні речовини, паприклад, диплацин, що утворюють стійкі з’єднання з Н-холінорецепторами та не дають змогиацетилхоліну взаємодіяти з ними. Медіатор ацетилхолін взаємодіє з рецепторами протягом короткого – часу, після чого комплекс медіатор-циторецептор розпадається і медіатор руйнується ферментом ацетилхолінестеразою, активність якого в синапсі дуже висока.
6) Механізми скорочення та розслаблення скелетних м'язів
 Термін “спряження збудження із скороченням” означає взаєзв’язок збудження в скелетних м’язах (виникнення та поширення ПД по мембрані волокна) та його скорочення, тобто актоміозинової взаємодії.
Термін “спряження збудження із скороченням” означає взаєзв’язок збудження в скелетних м’язах (виникнення та поширення ПД по мембрані волокна) та його скорочення, тобто актоміозинової взаємодії.
В стані спокою взаємодії актинових та міозинових протофібрил немає тому, що активні центри актину заблоковані регуляторними білками тропоніном та тропоміозином. Знімають блокаду іони Са2+, концентрація яких в саркоплазмі в стані спокою низька 10–8 ммоль/л.
ПД, що виникають в постсинаптичній мембрані поширюються вздовж всієї довжини мембрани м’язового волокна, у тому числі і по мембрані Т-трубочок (це вгинання зовнішньої мембрани м’язових клітин всередину волокна).
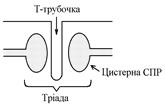 Безпосередньо поруч з Т-трубочками розташовані цистерни саркоплазматичного ретикулуму (СПР – частина аграпулярної епдоплазматичної сітки). Т-трубочки з двома поруч розташованими цистернами утворюють тріади. СПР крім цистернмаютьпоздовжні трубочки, в яких локалізуються кальцієві насоси – вони активно транспортують іони – Са2+із саркоплазми в СПР і в його цистернах накопичується велика кількість іонів Са2+. При русі ПД по мембрані Т-трубочок в мембрані цистерн СПР відкриваються кальцієві канали Ù іони Са2+ по градієнту концентрації виходять з цистерн СПР у саркоплазму Ù підвищення концентрації іонів Са2+ в саркоплазмі міоцита з 10–8 до 10–5 ммоль/лÙ дифузія іонів Са2+ до протофібрилÙ взаємодія з регуляторним білком тропоніномÙ зміна третинної конформації тропоніну та тропоміозину Ù відкриття активних центрів актину Ù взаємодія головок міозину з активними центрами актину (м’язове скорочення).
Безпосередньо поруч з Т-трубочками розташовані цистерни саркоплазматичного ретикулуму (СПР – частина аграпулярної епдоплазматичної сітки). Т-трубочки з двома поруч розташованими цистернами утворюють тріади. СПР крім цистернмаютьпоздовжні трубочки, в яких локалізуються кальцієві насоси – вони активно транспортують іони – Са2+із саркоплазми в СПР і в його цистернах накопичується велика кількість іонів Са2+. При русі ПД по мембрані Т-трубочок в мембрані цистерн СПР відкриваються кальцієві канали Ù іони Са2+ по градієнту концентрації виходять з цистерн СПР у саркоплазму Ù підвищення концентрації іонів Са2+ в саркоплазмі міоцита з 10–8 до 10–5 ммоль/лÙ дифузія іонів Са2+ до протофібрилÙ взаємодія з регуляторним білком тропоніномÙ зміна третинної конформації тропоніну та тропоміозину Ù відкриття активних центрів актину Ù взаємодія головок міозину з активними центрами актину (м’язове скорочення).
В основі м’язового скорочення лежить ковзання актинових протофібрил відносно міозинових “теорія ковзання”.
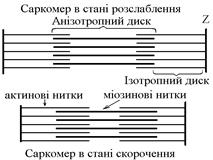 Саркомер – сруктурно-функціональний елемент скорочувального апарату скелетних м’язів. Вони утворені пучками міофібрил, які відокремлені один від одного перпендикулярними смугами – Z-лініями. До Z-ліній прикріпляються одним своїм кінцем тонкі актинові нитки. Інші кінці актинових ниток спрямовані до центру саркомера і входять у проміжки між товстими міозиновими нитками. Частина саркомера, яка примикає до Z лінії та утворена тільки актиновими протофібрилами, має назву І-дисків (ізотропних); слідом за ними розташовані А-диски (анізотропні) – частина саркомера, де має місце взаємне перекриття актинових та міозинових протофібрил. При вкороченні м’яза, в, ході його скорочення, коротшає довжина протофібрил Ù вкорочення довжини кожного саркомера. Але при цьому довжина анізотропних дисків не зменшується, а зменшується довжина ізотропних дисків. Це є наслідком ковзання актинових протофібрил відносно міозинових в напрямку до центру саркомера.
Саркомер – сруктурно-функціональний елемент скорочувального апарату скелетних м’язів. Вони утворені пучками міофібрил, які відокремлені один від одного перпендикулярними смугами – Z-лініями. До Z-ліній прикріпляються одним своїм кінцем тонкі актинові нитки. Інші кінці актинових ниток спрямовані до центру саркомера і входять у проміжки між товстими міозиновими нитками. Частина саркомера, яка примикає до Z лінії та утворена тільки актиновими протофібрилами, має назву І-дисків (ізотропних); слідом за ними розташовані А-диски (анізотропні) – частина саркомера, де має місце взаємне перекриття актинових та міозинових протофібрил. При вкороченні м’яза, в, ході його скорочення, коротшає довжина протофібрил Ù вкорочення довжини кожного саркомера. Але при цьому довжина анізотропних дисків не зменшується, а зменшується довжина ізотропних дисків. Це є наслідком ковзання актинових протофібрил відносно міозинових в напрямку до центру саркомера.
 Причиною руху актинових ниток відносно міозинових (їх ковзання) є “нахил” голівки після її приєднання до активного центру актину. Велика кількість голівок, що входять до складу актоміозинових містків, одночасно роблять “нахил” Ù нитка актину переміщується до центру саркомера вздовж нитки міозину. За рахунок одного такого “нахилу” саркомер (м’яз) може вкоротитися на 1% своєї вихідної довжини. Ступінь вкорочення м’яза може досягати 50% Ù такі “нахили” голівок під час одного м’язового скорочення мають повторитися 50 разів. Це можливо, якщо актоміозинові містки після “нахилу” розпадаються (дисоціюють) Ù голівка міозину взаємодіє з наступним активним центром актину Ù наступний “нахил” Ù розпад містку і т.д. На “нахил” однієї головки міозину витрачається енергія однієї молекули АТФ. Для того, щоб відбувся розпад актоміозинового містка необхідно, щоб до голівки міозину приєдналася молекула АТФ (але енергія АТФ при цьому не витрачається).
Причиною руху актинових ниток відносно міозинових (їх ковзання) є “нахил” голівки після її приєднання до активного центру актину. Велика кількість голівок, що входять до складу актоміозинових містків, одночасно роблять “нахил” Ù нитка актину переміщується до центру саркомера вздовж нитки міозину. За рахунок одного такого “нахилу” саркомер (м’яз) може вкоротитися на 1% своєї вихідної довжини. Ступінь вкорочення м’яза може досягати 50% Ù такі “нахили” голівок під час одного м’язового скорочення мають повторитися 50 разів. Це можливо, якщо актоміозинові містки після “нахилу” розпадаються (дисоціюють) Ù голівка міозину взаємодіє з наступним активним центром актину Ù наступний “нахил” Ù розпад містку і т.д. На “нахил” однієї головки міозину витрачається енергія однієї молекули АТФ. Для того, щоб відбувся розпад актоміозинового містка необхідно, щоб до голівки міозину приєдналася молекула АТФ (але енергія АТФ при цьому не витрачається).
Для розслаблення скелетного м’язу необхідно, щоб зменшилась концентрація іонів Са2+в саркоплазмі. Це досягається завдяки дії кальцієвих насосів повздовжніх трубочок СПР Ù активний транспорт іонів Са2+ в СПР Ù зменшення його концентрації в саркоплазмі до рівня 10–8 ммоль/л Ù від’єднання іонів Са2+ від регуляторного білка тропоніну Ù відновлення вихідної третинної структури тропоніну та тропоміозину Ù блокада активних центрів актину Ù розслаблення м’язу.
Таким чином, під час м’язового скорочення енергія АТФ витрачається:
1. На роботу натрій-калієвого насосу зовнішньої мембрани м’язового волокна (цей насос підтримує градієнт концентрації іонів К+ та Na+ поза та всередині клітини, що необхідно для підтримки на необхідному рівні ПС та для розвитку ПД на мембрані).
2. На “нахил” головки міозину.
3. На роботу кальцієвого насосу СПР Ù розслаблення м’язу.
7) Види м’язових скорочень: одинокі та тетанічні; ізотонічні та ізометричні.
В залежності від режимів навантаження виділяють наступні види м’язового скорочення.
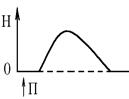 1. Ізометричного скорочення – скорочення при незмінній довжинні м’яза.Довжина м’яза при скороченні залишається сталою, але змінюється її напруження (Н). В експерименті таке скорочення можна отримати, якщо ізольований м’яз закріпити з двох сторін та стимулювати електричним струмом. В умовах цілісного організму ізометричне скорочення буває, коли людина намагається, але не може підняти вантаж.
1. Ізометричного скорочення – скорочення при незмінній довжинні м’яза.Довжина м’яза при скороченні залишається сталою, але змінюється її напруження (Н). В експерименті таке скорочення можна отримати, якщо ізольований м’яз закріпити з двох сторін та стимулювати електричним струмом. В умовах цілісного організму ізометричне скорочення буває, коли людина намагається, але не може підняти вантаж.
 2. Ізотонічне скорочення – скорочення при незмінному напруженні м’яза. Напруження м’яза при його скороченні залишається сталим, а довжина змінюється (Δ l). В експерименті таке скорочення можна отримати, якщо один кінець ізольованого м’яза закріпити, а інший залишити вільним та стимулювати. В умовах цілісного організму скорочення м’язів максимально наближується до ізотонічного, якщо відбувається згинання ненавантажених кінцівок.
2. Ізотонічне скорочення – скорочення при незмінному напруженні м’яза. Напруження м’яза при його скороченні залишається сталим, а довжина змінюється (Δ l). В експерименті таке скорочення можна отримати, якщо один кінець ізольованого м’яза закріпити, а інший залишити вільним та стимулювати. В умовах цілісного організму скорочення м’язів максимально наближується до ізотонічного, якщо відбувається згинання ненавантажених кінцівок.
3. Змішане (ауксотонічне) скорочення – скорочення, при якому змінюється довжина та напруження м’яза. Такий тип м’язових скорочень найчастіше зустрічається в умовах цілісного організму.
В залежності від частоти стимуляції виділяють наступні види м’язового скорочення:
1. Одиночне – спостерігається у відповідь на одиничний стимул чи на їх серію (якщо інтервал між подразниками більший чи рівний тривалості одиночного м’язового скорочення). Виділяють такі періоди м’язового скорочення:
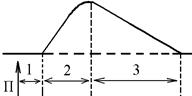 - латентий період (1) – інтервал часу від дії подразника (П) до початку скорочення;
- латентий період (1) – інтервал часу від дії подразника (П) до початку скорочення;
- період скорочення (2) – під час цього періоду довжина м’яза зменшується;
- період розслаблення (3) – під час цього періоду м’яз розслаблюється, його довжина збільшуєтся до вихідного рівня.
2. Тетанічне – спостерігається в умовах сумації одиночних м’язових скорочень (одне скорочення накладається на інше). Така сумація можлива, якщо м’яз подразнюється серією стимулів і кожен наступний стимул діє тоді, коли попереднє одиночне скорочення ще не закінчилось (тобто, інтервал між стимулами має бути меншим тривалості одиночного м’язового скорочення).
Виділяють наступні види тетанічних скорочень:
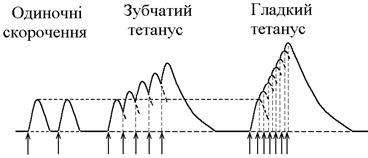
Зубчастий тетанус – виникає на базі одиночних скорочень, якщо кожен наступний стимул попадає на період розслаблення одиночого м’язового скорочення. Отже, інтервал між стимулами в серії має бути менший тривалості одиночного м’язового скорочення, але більший, чим cума тривалості латентного періоду і періоду вкорочення одиночного скорочення.
Гладкий тетанус – виникає на базі одиночних скорочень, якщо кожен наступний стимул попадає на латентний період або період вкорочення одиночного м’язового скорочення. Отже, інтервал між стимулами в серії повинний бути меншимчи рівним сумі тривалості латентного періоду одиночного скорочення, але більше тривалості латентного періоду одиночного скорочення.
Нижче зображений запис реєстрації скорочень м’яза при збільшенні частоти її стимуляції. Видно, що при збільшенні частоти стимуляції на фазі одиночних скорочень виникають тетанічні, спочатку зубчастий, а потім гладкий тетанус. Тетанічні скорочення відрізняється від одиночного наступними параметрами:
1. Тривалість тетанічного скорочення більша, ніж одиночного.
2. Амплітуда (сила) тетанічного скорочення більша, ніж одиночного. Амплітуда гладкого тетануса більша, ніж зубчастого.