Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Соотносительные преобразования органов
|
|
Рассмотренные выше филогенетические преобразования отдельных биологических структур протекают в эволюционирующих группах организмов как целостных системах. Устойчивые взаимозависимости органов и систем, проявляющиеся в филогенезе, называют координациями.
Механизмы соотносительных преобразований биологических структур различны, в связи с чем выделяют три их группы: биологические, динамические и топографические.
Биологические координации наблюдаются между структурами, непосредственно не связанными ни по функциям, ни по месту положения. Основным связующим звеном между ними являются адаптации к определенным условиям обитания. Так, у большинства эндопаразитов (см. § 18.6) сильно развиты половая система и органы прикрепления к телу хозяина, но при этом недоразвиты органы чувств и опорно-двигательный аппарат. Млекопитающие, обитающие на деревьях, обычно имеют стереоскопическое зрение и сильно развитый мозжечок. Позвоночные, которые дышат кислородом, растворенным в воде, имеют хвостовой плавник, жабры, слабо развитый передний мозг и содержат гемоглобин, способный связываться с кислородом при низком его парциальном давлении в среде. Все позвоночные, имеющие матку, обладают также совершенной системой терморегуляции.
Динамические координации выражаются во взаимном соответствии структур, связанных функционально. Тесные динамические координации имеются, например, между органами кровеносной и дыхательной систем. Так, животные, дышащие легкими, имеют трех- или четырехкамерное сердце и два круга кровообращения. Степень развитости нервных центров всегда соответствует интенсивности функционирования иннервируемых органов. Например, строение органа обоняния и обонятельные доли мозга у млекопитающих высоко развиты, в то время как у птиц примитивное строение периферической части обонятельного анализатора соответствует малым размерам обонятельных долей мозга. Это связано с тем, что в ориентации млекопитающих обоняние играет первостепенную роль, а для птиц оно не имеет большого значения.
Топографические координации проявляются между структурами, связанными друг с другом пространственно. Так, для каждого типа животного царства характерен своеобразный общий план строения, выражающийся в определенном взаимном расположении основных органов и систем. Например, у всех представителей типа Хордовые на спинной стороне тела расположена нервная трубка, под ней лежат хорда, пищеварительная трубка и брюшной кровеносный сосуд, а по бокам тела — производные мезодермы (см. разд. 13.5.4).
Все типы координации характеризуются высокой степенью устойчивости. Так, хордовые животные, известные с конца протерозойской эры, сохранили неизменными общий план строения до настоящего времени, на протяжении более 500 млн. лет. Феномен паразитизма возник значительно раньше появления хордовых, и поэтому комплекс адаптации к паразитическому образу жизни (см. § 18.6) является еще более древним. Длительно существуют и другие, более частные координации, возникающие вместе с появлением новых таксономических или экологических групп организмов.
Высокая устойчивость филогенетических координации обеспечивается целостностью онтогенеза каждой конкретной особи, развитие всех биологических структур которой протекает в строгом взаимном соответствии. Такое соответствие структур развивающегося организма в онтогенезе называют онтогенетическими корреляциями. Различают геномные, эргонтические и морфогенетические корреляции.
Геномные корреляции обеспечиваются целостностью генетической конституции развивающегося организма. Ведущими механизмами геномных корреляций являются генный баланс генотипа, сцепленное наследование генов, различные формы взаимодействия генов, а также плейотропность. Так, генные системы, регулирующие процессы пролиферации и избирательной гибели клеток на различных этапах органогенеза (см. разд, 8.2.1, 8.2.4), приводят к аллометрическому росту органов (см. разд. 8.3.3), благодаря чему появляются, например, удлиненный клюв, шея и задние конечности у большинства болотных птиц, длинная шея и ноги у жирафа, а также отличающиеся друг от друга пропорции тела у мужчин и женщин.
Морфогенетические корреляции возникают между органами, пространственно связанными между собой. Они основаны либо на феномене эмбриональной индукции (см. разд. 8.2.6), либо на общности эмбриональных закладок органов. Эти корреляции проявляются уже на ранних стадиях онтогенеза, когда еще отсутствуют функциональные связи между формирующимися органами. Так, зачаток хорды обусловливает развитие нервной трубки на спинной стороне зародыша и дифференцировку скелетогенной ткани внутренних частей сомита — склеротома в хрящ или кость, а глазной бокал (вырост переднего мозга) — формирование хрусталика при морфогенезе глаза.
Примером развития ряда структур из одного общего зачатка является формирование у млекопитающих и человека из закладок 1-й и 2-й пар жаберных дуг первичных челюстей, подъязычной кости, части хрящей гортани, шиловидного отростка черепа и трех слуховых косточек. При нарушении развития закладок жаберных дуг обычно образуется комплекс аномалий указанных структур, называемый аномаладом первых жаберных дуг.
Эргонтические корреляции обусловлены функциональной взаимозависимостью органов и частей организма. Они возникают на более поздних стадиях развития, когда органы начинают функционировать, и проявляются, например, в соответствии между степенью развитости мышцы, костных выступов, к которым она прикрепляется, и интенсивностью ее кровоснабжения. К такого рода корреляциям относят также соответствие вторичных половых признаков развитию гонад.
Ведущие корреляции в онтогенезе — геномные. В конечном счете именно они лежат в основе других корреляций, значение которых на протяжении онтогенеза меняется. Это связано с первичностью изменений генотипа в процессе филогенеза.
Система корреляций и сама подвергается эволюционным преобразованиям. В процессе эволюции видоизменяются вначале более частные корреляции, в то время как наиболее общие могут воспроизводиться в конкретных онтогенезах очень длительное время. В результате в ходе исторического развития происходит как отбор наиболее общих корреляций, имеющих значение при любых перестройках организма и в различных условиях обитания (общий план строения, соответствие между степенью развитости нервного центра и иннервируемыми органами), так и накопление локальных корреляций частного значения, формирующихся у организмов разных видов и отражающих специфику их образа жизни.
Корреляции общего значения обусловливают преемственность формообразовательных процессов в череде поколений организмов данного типа организации, а частные корреляции — многообразие конкретных форм жизни.
Между филогенетическими координациями и онтогенетическими корреляциями имеется теснейшая связь. Очевидно, что корреляции существуют и воспроизводятся в поколениях благодаря тому, что на протяжении предшествующей эволюции органов они преобразовывались скоординированно. С другой стороны, филогенетические координации в последующей эволюции организмов будут реализовываться благодаря воспроизведению онтогенетических корреляций в ходе индивидуального развития конкретных особей. Таким образом, в виде соотношения корреляций и координации проявляется диалектическое единство онто- и филогенеза как целостного процесса исторического развития живого.
Сопоставление конкретных форм соотносительных преобразований органов приводит к выводу о том, что морфогенетические корреляции и топографические координации точно так же, как эргонтические корреляции и динамические координации, взаимно обусловливают друг друга. В паре геномные корреляции — биологические координации также обнаруживается соответствие.
Осознание целостности и взаимообусловленности индивидуального и исторического развития необходимо врачу в связи с тем, что соотносительное преобразование биологических структур лежит в основе нормального развития организма человека, а нарушение даже одного незначительного компонента развивающейся системы может повлечь за собой возникновение целого комплекса нарушений в других ее элементах. С этим связано то, что практически все врожденные пороки развития, а также многочисленные генные болезни характеризуются не отдельными патологическими признаками, а синдромами, представляющими собой комплексы симптомов, связанных между собой процессом формирования.
Сердце у всех позвоночных закладывается на ранних этапах развития кпереди от глотки под челюстной дугой. В его морфогенезе участвует глотка как эмбриональный индуктор. Если это свойство глотки нарушено, тсгсердце может задержаться на двух- и трехкамерном уровне развития, при этом может быть нарушено и его перемещение в загрудинную область — шейная эктопия сердца (см. § 14.4). Эти явления — результат нарушений морфогенетических корреляций в развитии шейной области. Часто этот порок развития сопровождается нарушением отходящих от сердца сосудов (персистирование общего эмбрионального ствола, двух дуг аорты и т.д.) и недоразвитостью легких.
В возникновении данных аномалий ведущим механизмом выступает нарушение эргонтических корреляций сердце — сосуды — легкие. Первичным нарушением в описанном комплексе признаков является, вероятно, нарушение генетического контроля эмбриональной индукции, описанное ранее (см. разд. 8.2.6). Таким образом, приведенный пример иллюстрирует взаимоотношение разных форм соотносительных преобразований органов при формировании сложного комплекса патологических признаков, имеющих в целом атавистическую природу.
Примером нарушения чисто геномных корреляций является синдром Дауна. Увеличение доз генов 21-й хромосомы или ее части приводит к формированию тяжелой умственной отсталости, ослаблению тонуса мышц, аномалиям мозгового черепа и мягких частей лица, светлой пигментации волос и глаз.
Кроме филогенетических координации, подкрепляемых в каждом поколении онтогенетическими корреляциями, целостность развивающегося организма отражают и такие соотносительные преобразования органов, как субституция и гетеробатмия.
Субституция — это такое эволюционное преобразование, при котором один орган замещается другим, выполняющим обычно ту же функцию с большей интенсивностью. При этом наблюдается развитие этих органов в разных направлениях. Один обычно подвергается редукции, другой — эволюционирует прогрессивно. Так, хорда замещается позвоночником и превращается в рудиментарное образование, а первичные хрящевые челюсти позвоночных заменяются вторичными костными (см. разд. 14.2.1). Это примеры гомотопной субституции, когда новый орган возникает на месте старого. При гетеротопной субституции заменяющий орган находится на новом месте. Так, функцию печени как органа кроветворения берет на себя красный костный мозг. Выделительная функция выполняется у рыб и земноводных туловищной почкой, а у пресмыкающихся и млекопитающих — тазовой.
Гетеробатмия — это такое эволюционное преобразование, при котором в одной группе организмов обнаруживается разный уровень эволюционной продвинутое™ и специализации разных частей одного и того органа, разных органов одной и той же системы или разных частей организма. Примером может являться человек, головной мозг которого за короткое время антропогенеза претерпел колоссальные морфофизиологические изменения, в то время как пищеварительная система соответствует уровню развития других приматов.
Гетеробатмия, наблюдающаяся внутри одной и той же системы органов в разных филогенетических группах, обусловливает феномен компенсации функций, благодаря которому одни и те же экологические задачи решаются разными способами. Так, грызуны и копытные млекопитающие питаются одинаковой растительной пищей, но у первых наиболее выраженные адаптации к растительноядности проявляются в строении зубов и морфофизиологии слюнных желез, в то время как вторые на фоне примитивной зубной системы имеют высокоспециализированные желудок и кишечник. Явления гетеробатмии и, следовательно, компенсации функций имеют огромное эволюционное значение в связи с тем, что в организме, даже вступившем на путь узкой специализации, всегда остаются органы и системы относительно мало специализированные, которые при меняющихся условиях могут еще прогрессивно развиваться, раскрывая перед такими филогенетическими группами новые адаптивные возможности.
96.Филогенез наружных покровов хордовых животных. Онтофилогенетические пороки развития наружных покровов у человека.
Покровы любых животных всегда выполняют функцию восприятия внешних раздражении, а также защищают тело от вредных воздействий среды. Интенсификация первой функции покровов приводит в процессе эволюции многоклеточных животных к возникновению нервной системы и органов чувств. Интенсификация второй функции сопровождается дифференцировкой. Характерно также расширение функций, в результате чего кожа как орган защиты участвует также в газообмене, терморегуляции и выделении, выкармливании потомства. Это связано с усложнением строения слоев кожи, появлением и дальнейшим преобразованием многочисленных придатков и желез.
У всех хордовых кожа имеет двойное — экто- и мезодермальное — происхождение. Из эктодермы развивается эпидермис, из мезодермы — дерма. Для бесчерепных характерна слабая степень дифференцированности обоих слоев кожи. Эпидермис однослойный цилиндрический, содержащий одноклеточные слизистые железы, дерма рыхлая, содержит небольшое количество соединительнотканных клеток.
В подтипе Позвоночные эпидермис становится многослойным, причем в нижнем слое клетки постоянно размножаются, а в верхних слоях — дифференцируются, гибнут и слущиваются. В дерме появляются соединительнотканные волокна, придающие покровам прочность. Кожа образует придатки, многообразные в зависимости от образа жизни и уровня организации, а также железы, выполняющие различные функции.
У рыб в эпидермисе железы одноклеточные. Как и у ланцетника, они выделяют слизь, облегчающую движения в воде. Тело рыб покрыто чешуей, имеющей разное строение в зависимости от их систематического положения. Чешую хрящевых рыб называют плакоидной. Она имеет форму шипа и состоит из дентина, покрытого снаружи эмалью (рис. 14.1). Дентин мезодермального происхождения, он образуется за счет функционирования соединительнотканных клеток, выпячивающихся снаружи в виде сосочка. Эмаль, представляющая собой более твердое, чем дентин, неклеточное вещество, формируется сосочком эпидермиса и покрывает плакоидную чешую снаружи.
Вся поверхность тела хрящевых рыб, а также ротовая полость, слизистая оболочка которой происходит из эктодермы, покрыты плакоидной чешуей. Естественно, что функции чешуи в ротовой полости связаны с захватом и удержанием пищи, поэтому они сильно увеличены и являются зубами. У костных рыб чешуя иного типа. Она имеет вид тонких круглых костных пластинок, покрытых тонким слоем эпидермиса. Костная чешуя развивается полностью за счет дермы, но по происхождению связана с примитивной плакоидной.
Кожа примитивных вымерших земноводных — стегоцефалов — соответствовала покровам рыб и тоже была покрыта чешуей. Современные амфибии имеют тонкую гладкую кожу без чешуи, принимающую участие в газообмене. Этому способствует наличие большого количества многоклеточных слизистых желез, секрет которых постоянно увлажняет покровы и обладает бактерицидными свойствами. Некоторые кожные железы ряда земноводных дифференцировались в органы-продуценты токсинов, защищающих их от врагов (см. разд. 23.1).
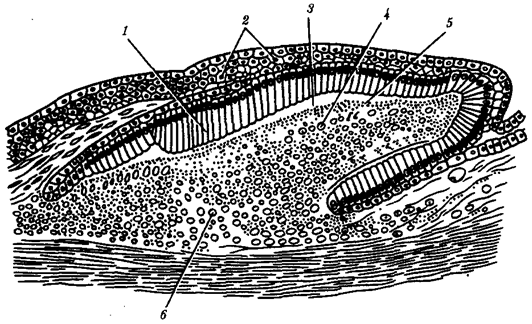
Рис. 14.1. Закладка плакоидной чешуи:
1 —клетки-эмалеобразователи, 2— эпидермис, 3— эмаль, 4— склеробласты—дентинообразователи, 5— дентин, 6— сосочек дермы
Пресмыкающиеся, перешедшие полностью к наземному существованию, имеют сухую кожу, не участвующую в дыхании. Верхний слой эпидермиса ороговевает. Роговые чешуйки у некоторых рептилий тонки и эластичны, у других—сливаются вместе, образуя, как у черепах, мощный роговой панцирь. Большинство пресмыкающихся по мере роста линяют, сбрасывая периодически свой роговой покров. Современные пресмыкающиеся не имеют кожных желез.
Кожные покровы млекопитающих построены наиболее сложно в связи с выполнением ими многообразных функций. Характерны различные производные кожи: волосы, когти, рога, копыта, а также потовые, сальные и млечные железы. Более примитивные млекопитающие — насекомоядные, грызуны и некоторые другие — наряду с волосяным покровом сохранили также роговые чешуи на хвосте. Волосы у них растут в промежутках между чешуйками, группами по 3—7. У более прогрессивных млекопитающих, утративших чешуи, сохраняется такое же расположение волос (рис. 14.2), покрывающих практически все тело, кроме некоторых участков, например подошв и ладоней у человека.
Волосы многих млекопитающих дифференцированы на типичные, служащие для терморегуляции, и большие, или вибриссы, основания которых связаны с чувствительными нервными окончаниями. У большинства млекопитающих вибриссы расположены в области рта и носа, у приматов они редуцированы в связи с усилением осязательной функции передних конечностей, у многих яйцекладущих и сумчатых — разбросаны по всему телу. Этот факт может свидетельствовать о том, что волосяной покров предков млекопитающих первично выполнял осязательные функции, а затем, по мере увеличения количества волос, стал принимать участие в терморегуляции. В онтогенезе человека закладывается большее количество волосяных зачатков, но к концу эмбриогенеза наступает редукция большинства из них.
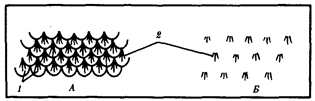 Рис. 14.2. Расположение волос у млекопитающих. А—на хвосте грызунов; Б— на других участках тела:
1 —роговые чешуйки, 2— группы волос, расположенные в шахматном порядке
Рис. 14.2. Расположение волос у млекопитающих. А—на хвосте грызунов; Б— на других участках тела:
1 —роговые чешуйки, 2— группы волос, расположенные в шахматном порядке
| 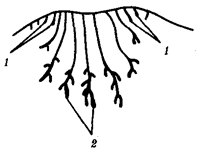 Рис. 14.3. Строение развивающегося соска млекопитающего
Виден постепенный переход от потовых (1) к млечным (2) железам
Рис. 14.3. Строение развивающегося соска млекопитающего
Виден постепенный переход от потовых (1) к млечным (2) железам
|
Потовые железы млекопитающих гомологичны кожным железам амфибий. Их секрет может быть слизистым, содержать белки и жир. Некоторые потовые железы дифференцировались у ранних млекопитающих в млечные железы. У яйцекладущих (утконос, ехидна) млечные железы сходны с потовыми по строению и развитию. По краям развивающегося соска молочной железы можно обнаружить последовательные переходы от типичных потовых к млечным железам (рис. 14.3). Количество млечных желез и сосков коррелирует с плодовитостью (от 25 до одной пары), но в эмбриогенезе всех млекопитающих на брюшной поверхности закладываются «млечные линии», тянущиеся от подмышечной впадины до паха. Впоследствии на этих линиях дифференцируются соски, большая часть которых затем подвергается редукции и исчезает. Так, в эмбриогенезе человека закладывается вначале пять пар сосков, а впоследствии остается лишь одна (рис. 14.4).
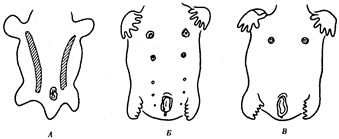
Рис. 14.4. Эмбриогенез передней брюшной стенки человека. А — зародыш в возрасте 5 недель (видны млечные линии); Б — дифференцировка пяти пар сосков; В — зародыш в возрасте 7 недель
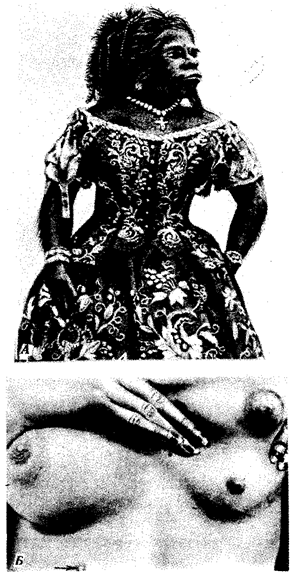
Рис. 14.5. Атавистические аномалии развития кожи.
А — гипертрихоз; Б — полимастия
Сальные железы образуются в коже только у млекопитающих. Их секрет, смазывая волосы и поверхность кожи, придает им несмачиваемость и эластичность.
Онтогенез покровов и придатков кожи млекопитающих и человека отражает их эволюцию по типу архаллаксиса. Действительно, ни зачатки роговых чешуи, характерных для пресмыкающихся, ни более ранние формы придатков кожи в их эмбриогенезе не рекапитулируют. При этом на стадии вторичного органогенеза развиваются сразу зачатки волосяных фолликулов. Нарушения раннего онтогенеза кожных покровов человека могут вызвать возникновение некоторых малосущественных атавистических пороков развития: гипертрихоз (повышенное оволосение), полителию (увеличенное количество сосков), полимастию (увеличенное количество млечных желез) (рис. 14.5). Все они связаны с нарушением редукции избыточного количества этих структур и отражают эволюционную связь человека с наиболее близкими предковыми формами — млекопитающими. Именно поэтому у человека и других млекопитающих невозможно рождение потомства с атавистическими признаками кожных покровов, характерными для более отдаленных предков. Одним из самых известных признаков недоношенности новорожденных является повышенное оволосение кожи. Вскоре после рождения избыточные волосы обычно выпадают, а их фолликулы редуцируются.
98.Филогенез пищеварительной системы хордовых. Онтофилогенетические пороки пищеварительной системы у человека.
Уникальной особенностью организации хордовых является филогенетическая, эмбриональная, а также функциональная связь пищеварительной и дыхательной систем. Действительно, только у хордовых дыхательная система развивается на базе пищеварительной и на первых этапах эволюции функционирует совместно с ней. Так, у ланцетника, сохранившего в значительной степени черты организации предковых форм, специализированных органов дыхания нет, а дыхательную функцию выполняет глотка — передняя часть пищеварительной трубки, пронизанная сквозными отверстиями — жаберными щелями, главной функцией которых является фильтрация воды. Челюстного аппарата у ланцетника нет, и активно питаться он не может. Поэтому источником его питания является взвесь органических частиц в воде, которые задерживаются жаберными перегородками, прилипая к слизи, обильно выделяемой железистыми клетками глотки. Постоянный ток свежей воды через глотку способствует газообмену в кровеносных сосудах, расположенных в жаберных перегородках. На капилляры эти сосуды не распадаются, что свидетельствует о второстепенности дыхательной функции глотки.
У более высокоорганизованных хордовых начиная с рыб пищеварительная и дыхательная функции осуществляются специализированными системами, объединенными анатомически общей полостью рта и глотки, а также развитием из общего энтодермального зачатка.
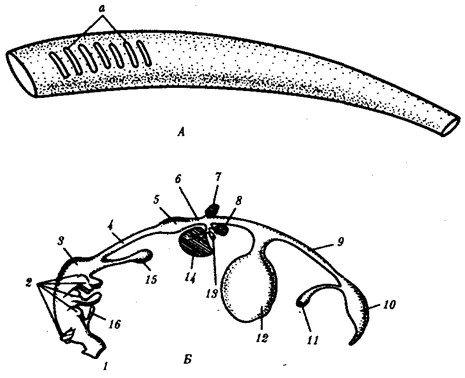
Рис. 14.18. Кишечная трубка позвоночных (А) и ее дифференцировка в эмбриогенезе человека (Б):
а— жаберные щели в глотке; 1 —ротовая полость, 2— 1—5-й глоточные карманы, 3— глотка, 4— пищевод, 5— желудок, 6— двенадцатиперстная кишка, 7—дорсальная поджелудочная железа, 8— вентральная поджелудочная железа, 9— тонкая кишка. 10 —клоака, 11 — мочевой пузырь, 12— желточный мешок, 13— желчный пузырь, 14— печень, 15— легкие, 16 —закладка щитовидной железы
Тесная связь обеих систем в филогенезе определяется в первую очередь их топографическими и динамическими координациями, а развитие в онтогенезе — морфогенетическими и эргонтическими корреляциями. Пищеварительная и дыхательная системы хордовых в эмбриогенезе закладываются вначале в виде прямой трубки, подразделяющейся на три участка. Переднюю ее часть, начинающуюся ротовым отверстием и заканчивающуюся переходом в глотку, называют stomodeum. Слизистая оболочка, выстилающая этот участок, эктодермального происхождения и в развитии связана с кожным эпидермисом и его производными (см. § 14.1). Средняя часть кишки начинается глоткой и заканчивается в том месте, где ее энтодермальная слизистая оболочка контактирует с эктодермальной слизистой оболочкой задней кишки, или proctodeum (рис. 14.18).