Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Синаптическая передача в нервной системе
|
|
Синапсы – это специализированная форма контакта между отростками нейронов и любыми возбудимыми образованиями – нейронами, мышечными или секреторными клетками. Термин «синапс» – от греческого synapsis – соприкосновение, соединение был введен Ч.Шеррингтоном в конце 19-го века. В головном и спинном мозге 60 – 80% всей нейрональной поверхности, включая сому, аксон и дендриты, занято синапсами. Каждый нейрон связан с тысячами других нервных клеток. Синапсы классифицируются по местоположению, характеру действия и способу передачи сигнала. По местоположению выделяют нервно-мышечные синапсы и нейро-нейрональные. Последние в свою очередь делятся на аксо-соматические, аксо-дендритные, аксо-аксональные и дендро-соматические. По характеру действия на воспринимающую структуру синапсы могут быть возбуждающими и тормозными. По способу передачи сигнала синапсы могут быть электрическими, химическими и смешанными.
Химические синапсы. В качестве примера можно рассмотреть аксо-соматический химический синапс. Аксон, подходя к телу другого нейрона, образует расширение, называемое пресинаптическим окончанием, или терминалью. Размеры пресинаптического окончания варьируют в диапазоне 0, 75 – 1, 5 мкм. Мембрана такого окончания называется пресинаптической. Под ней располагается синаптическая щель, ширина которой составляет 10 – 50 нм, а в нервно-мышечном синапсе 50 – 100 нм (для сравнения - расстояние между мембранами вне зоны их синаптического контакта не превышает 10 – 15 нм). Содержимое синаптической щели напоминает гель. За синаптической щелью находится мембрана тела нейрона, которая в области синапса называется постсинаптической. В составе постисинаптической мембраны обычно выделяют субсинаптическую мембрану, расположенную в проекции пресинаптических структур. Субсинаптическая мембрана существенно отличается от остальных участков субсинаптической мембраны: она не содержит потенциалзависимых ионных каналов, а содержит только хемиозависимые ионные каналы (лигандчувствительные), открывающиеся под действием нейромедиатора. Наконец, на субсинаптической мембране может находиться специальный фермент, расщепляющий нейромедиатор, что способствует освобождению постсинаптического рецептора.
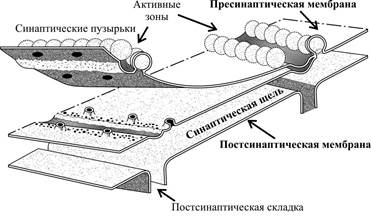
Рис. Ультраструктура химического синапса.
Медиатор содержится в пресинаптическом окончании в виде пузырьков (везикул). Примеры медиаторов синаптического возбуждения и торможения в ЦНС: ацетилхолин; катехоламины (адреналин, норадреналин, дофамин); серотонин; аминокислоты (глутаминовая, аспарагиновая, глицин, гамма-аминомаслянная кислота – ГАМК), полипептиды (вещество Р, энкефалин, соматостатин и др. Другие вещества – АТФ, гистамин, простогландины. Принято обозначать нейроны по тому медиатору, который выделяют его окончания – холинергические, адренергические, ГАМК–ергические и др. Установлено, что из пресинаптического окончания наряду с основным медиатором могут выделяться и другие биологически активные вещества – сомедиаторы. Например, вместе с ацетилхолином в холинергических синапсах могут высвобождаться АТФ, вазоактивный интестинальный пептид (ВИП), простагландины.
Синапсы и их рецепторы принято делить на ионотропные и метаботропные. В ионотропных синапсах медиаторы взаимодействуют с постсинаптическими рецепторами и способствуют открыванию ионного канала. В метаботропных нейромедиаторы влияют на специфические внутриклеточные биохимические процессы.
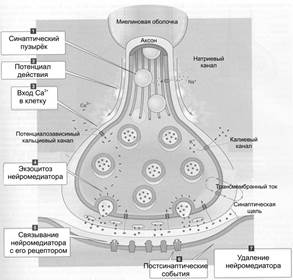
Рис. Синаптическая передача.
Под влиянием поступающих по пресинаптическому волокну нервных импульсов происходит деполяризация пресинаптической мембраны. Это приводит к открытию потенциалзависимых кальциевых каналов и входом кальция в пресинаптическое окончание. Ионы кальция способствуют слиянию мембраны синаптических пузырьков с пресинаптической мембраной. Выделение медиатора в синаптическую щель происходит не отдельными молекулами, а квантами, состоящими примерно из одинакового числа молекул, порядка нескольких тысяч. Выделившиеся кванты (порции) медиатора диффундируют через синаптическую щель к постсинаптической мембране, где связываются со специальными клеточными рецепторами, специфичными для каждого медиатора. Образовавшейся на постсинаптической мембране комплекс «медиатор – рецептор» активирует мембранные каналы, что повышает проницаемость мембраны для ионов и меняет ее потенциал покоя. Одномоментное формирование множества комплексов «медиатор – рецептор» приводит к генерации постсинаптического потенциала (ПСП).
Возбуждающие медиаторы вызывают появление на постсинаптической мембране возбуждающего постсинаптического потенциала (ВПСП). Его формирование обусловлено тем, что комплекс «медиатор – рецептор» активирует натриевые каналы мембраны и вызывает деполяризацию мембраны за счет поступления натрия внутрь клетки.

Рис. Возбуждающие и тормозные постсинаптические потенциалы (ВПСП и ТПСП соответственно) и токи (ВПСТ и ТПСТ). При взаимном наложении ВПСП и ТПСП происходит их суммация, однако результирующая деполяризация оказывается меньше, чем сумма этих потенциалов
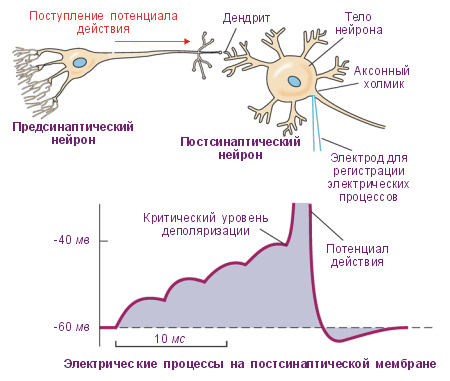
Амплитуда одиночного ВПСП довольно мала, от 0, 12 до 5 мВ и для уменьшения заряда мембраны до критического уровня необходима одновременная активация нескольких возбуждающих синапсов. ВПСП, образующиеся на постсинаптической мембране этих синапсов, способны суммироваться, приводя к росту амплитуды ВПСП. Процесс спонтанной деполяризации обычно возникает в аксонном холмике, где аксон еще не покрыт миелином и порог возбуждения низкий. Таким образом, ВПСП, возникающие в разных участках мембраны нейрона и на его дендритах, распространяются к аксонному холмику, где суммируются, деполяризуя мембрану до критического уровня и приводя к появлению потенциалов действия.
В тормозных синапсах выделяются другие медиаторы. Примером являются аминокислота глицин в тормозных синапсах спинного мозга и гамма-аминомаслянная кислота (ГАМК) – тормозной медиатор в нейронах головного мозга. Тормозной синапс может иметь тот же медиатор, что и возбуждающий, но иную природу рецепторов постсинаптической мембраны. Иначе говоря, эффект синаптической передачи – возбуждающий или тормозной определяется не столько природой медиатора, сколько постсинаптическими рецепторами.
Торможение связано с возникновением тормозного постсинаптического потенциала (ТПСП), обусловленного увеличением проницаемости мембраны для ионов калия. Калий выходит из клетки, в результате чего происходит гиперполяризация мембраны и снижение возбудимости. Аналогичный эффект во многих тормозных синапсах имеет активация каналов для хлора, увеличивающая его транспорт внутрь клетки. В результате возникновения ТПСП возбуждение нейрона становится невозможным.
Вся последовательность процессов, обеспечивающих синаптическую передачу с участием нейромедиатора, занимает 0, 2 – 0, 3 мс, это время носит название синаптической задержки. Удаление нейромедиатора из синаптической щели достигается его обратным захватом в пресинаптическое окончание или разрушением специфическими ферментами. Значительное число фармакологических препаратов, химических соединений, многие яды животного и растительного происхождения изменяют синаптическую передачу.
Синаптическая передача имеет следующие закономерности функционирования:
· Односторонний характер проведения возбуждения (от пресинаптической мембраны к постсинаптической)
· Наличие химических передатчиков – медиаторов,
· Свойства синапсов определяются природой постсинаптических рецепторов,
· Наличие хемочувствительных рецепторуправляемых каналов в постсинаптической мембране,
· Квантовый (порционный) характер освобождения медиатора.
· Количество квантов медиатора пропорционально частоте приходящих к синапсу нервных импульсов,
· Эффективность синаптической передачи возрастает при частом использовании синапсов («эффект тренировки») из-за ускорения синтеза медиатора и увеличения концентрации ионизированного кальция в пресинаптической терминали и, напротив, падает при редком использовании или бездействии, что носит название функциональной пластичности синапсов.
· Длительная высокая частота импульсов возбуждения ослабляет или прекращает синаптическую передачу (эффект истощения).
· Синаптическая передача не подчиняется закону «все или ничего».
· Характеризуется синаптической задержкой,
· В синапсе происходит трансформация ритма возбуждающих импульсов – частота импульсов в нерве может быть меньше или больше частоты возбуждения постсинаптической мембраны. Происходит усиление сигнала;
· конвергенция (схождение) многих сигналов на одной постсинаптической клетке;
· пластичность передачи сигналов (обучение, память и т. д.).
· Лабильность синапса меньше, чем нерва.
· Из всех звеньев рефлекторной дуги синапсы наиболее утомляемы и чувствительны к ядам и недостатку кислорода.
· Увеличение длительности постсинаптического потенциала при подавлении действия ферментов, разрушающих медиатор.
Электрические синапсы морфологически представляют собой слияние или сближение участков мембран, расстояние между которыми колеблется от 1 мкм до 0, 15 нм. При этом синаптическая щель между мембранами не сплошная, а прерывается мостиками контакта. Эти мостики образуют повторяющуюся ячеистую структуру синапса. В участках слияния мембран находятся каналы, через которые клетки могут обмениваться молекулами веществ. Функции электрических синапсов заключается в обеспечении срочных реакций организма. Электрический синапс сравнительно мало утомляем, устойчив к изменениям внешней и внутренней среды. Эти качества, наряду с быстродействием, обеспечивают высокую надежность его работы.