Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Структурные основы замыкания временных условных связей
|
|
Для изучения структурной основы временной связи ведутся многочисленные исследования на системном и клеточном уровнях; используют электрофизиологические и биохимические показатели динамики функционального состояния нервных и глиальных клеток с учетом результатов раздражения или выключения различных структур мозга; привлекают данные клинических наблюдений.
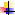 Удаление частей и всей коры больших полушарий. Исследования вопроса о локализации функций в коре в работах И.П. Павлова (1907–1909) показали широкое перекрытие проекционных зон, зависящее от природы сигналов и образуемых временных связей. И.П. Павлов (1927) выдвинул и обосновал представление о динамической локализации корковых функций. Если у взрослой собаки удалить затылочные области коры, то она утрачивает все сложные зрительные условные рефлексы и не может их восстановить. Но и в других областях коры находятся клетки, воспринимающие зрительные сигналы и способные их связывать с теми или иными действиями. Собака после удаления затылочных долей может вырабатывать лишь самые простые условные рефлексы, например, на вид освещенной фигуры. Заставить же ее различать две такие фигуры, сходные по форме, не удается.
Удаление частей и всей коры больших полушарий. Исследования вопроса о локализации функций в коре в работах И.П. Павлова (1907–1909) показали широкое перекрытие проекционных зон, зависящее от природы сигналов и образуемых временных связей. И.П. Павлов (1927) выдвинул и обосновал представление о динамической локализации корковых функций. Если у взрослой собаки удалить затылочные области коры, то она утрачивает все сложные зрительные условные рефлексы и не может их восстановить. Но и в других областях коры находятся клетки, воспринимающие зрительные сигналы и способные их связывать с теми или иными действиями. Собака после удаления затылочных долей может вырабатывать лишь самые простые условные рефлексы, например, на вид освещенной фигуры. Заставить же ее различать две такие фигуры, сходные по форме, не удается.
Возможность широкой взаимозаменяемести функций коры мозга в раннем онтогенезе соответствует свойствам малодифференцированной коры мозга млекопитающих в филогенезе. С этой точки зрения получают объяснение результаты опытов на крысах, у которых степень нарушения условных рефлексов оказалась зависящей не от конкретной области удаляемой коры, а от общего объема удаляемой корковой массы. На основании этих опытов было сделано заключение, что для условно-рефлекторной деятельности все отделы коры имеют одинаковое значение, кора «эквипотенциальна».
Убедительные доказательства дали опыты с временным обратимым выключением функций коры, которое проявляется в распространяющейся депрессии электрической активности при аппликации KCI на ее поверхность. При выключении таким образом коры мозга крысы и испытании в это время реакции животного на условные и безусловные раздражители можно видеть, что безусловные рефлексы полностью сохраняются, а условные — нарушаются.
Таким образом, результаты экспериментов с частичной и полной хирургической и функциональной декортикацией говорят о том, что у высших животных функции формирования точных и тонких условных рефлексов, способных обеспечить приспособительное поведение, в основном выполняет кора больших полушарий.
Корково-подкорковые отношения в процессах высшей нервной деятельности. Современные исследования подтверждают высказывание И.П. Павлова, что условно-рефлекторная деятельность осуществляется совместной работой коры и подкорковых структур. Взаимодействие коры и подкорковых структур мозга осуществляется и путем тонических влияний, регулирующих функциональное состояние нервных центров. Общеизвестно, как влияют настроение, эмоциональное состояние на эффективность умственной деятельности. И.П. Павлов говорил, что подкорка «заряжает» кору. Нейрофизиологические исследования механизмов подкорковых влияний на кору показали, что ретикулярная формация среднего мозга оказывает на нее восходящее активирующее действие. Получая коллатерали от всех афферентных путей, ретикулярная формация участвует во всех реакциях поведения, обусловливая деятельное состояние коры. Однако ее активирующее влияние при условном рефлексе организуется сигналами из проекционных зон коры (рис. 8). Раздражение ретикулярной формации вызывает изменение электроэнцефалограммы в виде ее десинхронизации, характерной для состояния активного бодрствования животных. Однако дальнейшие исследования, в частности опыты с разрушением и стимуляцией хвостатых ядер, и другие факты привели к заключению о наличии более сложных корково-подкорковых отношений.
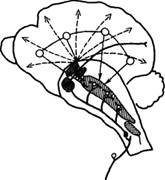
Рис. 8. Взаимодействие ретикулярной формации среднего мозга и коры (по Л.Г. Воронину):
жирными линиями обозначены афферентные специфические пути с коллатералями к ретикулярной формации, прерывистыми — восходящие пути к коре, тонкими — влияние коры на ретикулярную формацию, вертикальной штриховкой — облегчающая зона, горизонтальной — тормозная зона, клеточной штриховкой — таламические ядра.
Была сделана операция «расщепления» мозга путем перерезки мозолистого тела и передней комиссуры, а также продольного разделения перекреста зрительных нервов. После такой операции можно было вырабатывать разные условные рефлексы правого и левого полушарий, показывая разные фигуры правому или левому глазу. Если у оперированной таким образом обезьяны выработать условный рефлекс на световой стимул, подаваемый на один глаз, а затем подать его на другой глаз, то никакой реакции не последует. «Обучение» одного полушария оставляло другое «необученным». Однако при сохранении мозолистого тела и другое полушарие оказывается «обученным». М озолистое тело осуществляет межполушарный перенос навыка.
Безусловные рефлексы имеют центры на корковом уровне. И раздражитель, который сделан условным, также имеет представительство в коре. Отсюда возникло предположение, что временные связи условного рефлекса замыкаются между этими представительствами.