Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Экология популяций
|
|
Экологическая структура вида. Популяция как форма существования вида. Основные характеристики популяций.
Численность; критическая численность, эффективная численность. Плотность; оптимальная плотность. Пространственная структура популяции.
Рождаемость и смертность; абсолютные и относительные показатели рождаемости и смертности. Относительный прирост популяции. Абсолютный прирост популяции. Кривые изменения численности популяции. Жизненные стратегии.
Возрастная и половая структура популяции. Этологическая структура популяции; внутрипопуляционные группировки у животных.
В биологии существует множество концепций и множество определений вида. Одно из простейших определений вида гласит: вид – это совокупность организмов (особей), сходных между собой по ряду существенных признаков, населяющих определенный ареал, способных скрещиваться между собой и давать плодовитое потомство, похожее на родителей.
Ареал – это участок земной поверхности (территория или акватория), на котором существует и воспроизводит себя данный вид организмов. В большинстве случаев площадь ареала настолько велика, что организмы одного и того же вида должны адаптироваться к воздействию экологических факторов в разных условиях. Таким образом, вид обладает определенной экологической структурой.
Наиболее крупная внутривидовая экологическая группировка называется климатип, или географическая раса. С точки зрения систематики, климатип представляет собой подвид. Каждый климатип приспособлен к почвенно-климатическим условиям определенного географического региона.
Следующий уровень экологической структуры вида – экотип, или экологическая раса. С точки зрения систематики, экотип представляет собой разновидность. Каждый экотип приспособлен к условиям определенного местообитания. Например, экотип сосны на болоте отличается от экотипа сосны на песчаных дюнах.
Более дробная группировка – изореагент, или физиологическая раса. Это группа особей одного вида, реагирующих на действие экологических факторов сходным образом. С точки зрения систематики, изореагент представляет собой форму. Например, дуб черешчатый образует две формы (зимнюю и летнюю), у которых наблюдаются различные фотопериодические реакции.
И, наконец, мельчайшая внутривидовая экологическая группировка называется биотип. Биотип – это особи с одинаковым генотипом (линия, или клон), у которых совершенно одинаковые генетически обусловленные реакции на воздействие экологических факторов.
Основы популяционной биологии
Элементарной внутривидовой группировкой, которой соответствует собственная реализованная экологическая ниша, является популяция. В состав популяции могут входить разные экотипы, изореагенты и биотипы в различных соотношениях.
Наиболее полным и всеобъемлющим определением популяции является следующее:
Популяция – это минимальная самовоспроизводящаяся группировка особей одного вида, населяющая определенный ареал в течение длительного ряда поколений, образующая собственную генетическую систему, формирующая собственную экологическую нишу и более или менее изолированная от других подобных группировок данного вида.
Популяция есть форма существования вида и элементарная единица эволюции.
Практическое значение популяционной биологии связано с тем, что популяция есть единица эксплуатации, охраны и подавления.
Основные характеристики популяции
Популяции обладают рядом свойств, которые не присущи отдельно взятой особи или просто группе особей. К основным характеристикам популяции относятся: численность, плотность, рождаемость, смертность, относительный и абсолютный прирост.
1. Численность. Существует нижний предел численности, ниже которого популяция прекращает свое воспроизведение. Такая минимальная численность популяции называется критической. При определении критической численности нужно учитывать не всех особей, а только тех, которые принимают участие в размножении – это эффективная численность популяций.
Обычно численность популяций измеряется сотнями и тысячами особей. У человека минимальная численность популяций составляет около 100 особей. У крупных наземных млекопитающих численность популяций может снижаться до нескольких десятков особей (микропопуляции). У растений и беспозвоночных существуют также мегапопуляции, численность которых достигает миллионов особей.
В стабильных по численности популяциях число особей, оставляющих потомство, должно быть равно числу таких особей в предыдущих поколениях. Для управления численностью популяций необходимо знать их основные характеристики. Лишь в этом случае возможно прогнозирование изменения состояния популяции при воздействии на неё.
2. Плотность. Плотность популяции является производной характеристикой. Обычно плотность определяется как среднее число особей на единицу площади или объема занимаемого популяцией пространства. Плотность можно выражать также как массу (биомассу) членов популяции на единице площади или в единице объема.
Сравнительно многочисленные популяции могут обладать низкой плотностью. Низкая плотность снижает внутривидовую конкуренцию и повышает шансы отдельных членов популяции на выживание. При низкой плотности популяция не оказывает заметного влияния на функционирование сообществ и экосистем; например, энергия, затраченная хищником на поиск жертвы, не компенсируется энергетическими выгодами от съеденной жертвы – тогда становится бессмысленной пищевая специализация хищника.
Возможна и обратная ситуация: сравнительно малочисленная популяция может обладать высокой плотностью. В этом случае гарантируется образование брачных пар. При малочисленности, но высокой плотности популяция может оказывать значительное влияние на функционирование сообществ и экосистем.
Таким образом, низкая плотность популяции уменьшает ее шансы на воспроизведение, но увеличивает шансы на выживание. Высокая плотность, наоборот, увеличивает шансы на воспроизведение, но уменьшает шансы на выживание. Следовательно, каждая конкретная популяция должна обладать некоторой оптимальной плотностью.
Распределение плотности популяции тесно связано с ее пространственной структурой. Существует множество типов пространственной структуры популяций и, соответственно, типов популяционных ареалов: сплошные, разорванные, сетчатые, кольцевые, ленточные и комбинированные.
3. Рождаемость и смертность.
Рождаемость – это число новых особей, появляющихся в популяции за единицу времени. Новой особью (или особью нулевого возраста) может считаться зигота, яйцо, личинка или особь, вышедшая из-под родительской опеки. Различают абсолютную и относительную рождаемость.
Абсолютная рождаемость – это абсолютное число новых особей: например, в популяции в течение года родилось 156 новых особей.
Относительная (удельная) рождаемость – отношение числа новых особей к числу имевшихся особей; относительная рождаемость может рассчитываться или на одну особь, или на 1000 особей. Например, в популяции в начале года было 10 000 особей, а в течение года родилось 156 новых особей; тогда относительная рождаемость равна 156: 10000 = 0, 0156 на одну особь, или 0, 0156 · 1000 = 15, 6 на тысячу особей.
Существуют моноциклические (у растений монокарпические) виды, представители которых размножаются один раз в жизни, и полициклические (у растений поликарпические) виды, представители которых размножаются неоднократно.
Численность популяции может увеличиваться не только за счет рождаемости, но и за счет иммиграции особей из других популяций. Существуют зависимые и полузависимые популяции, которые поддерживают и увеличивают свою численность именно за счет иммиграции.
Смертность – это понятие, противоположное рождаемости. Различают абсолютную смертность (количество погибших особей за единицу времени) и относительную (удельную) смертность (количество погибших особей за единицу времени в расчете на одну особь или на 1000 особей).
В отличие от рождаемости смертность наблюдается постоянно. Характер смертности описывается таблицами и кривыми выживаемости, которые показывают, какая часть новорожденных особей дожила до определенного возраста.
Численность популяции может уменьшаться не только за счет смертности, но и за счет эмиграции особей.
4. Относительный прирост популяции. Этот параметр представляет собой разность между относительной рождаемостью и относительной смертностью. Это важнейшая характеристика популяции (обычно обозначается символом r). Относительный прирост популяции может быть положительным, нулевым и отрицательным.
Для изолированной популяции: прирост = рождаемость – смертность. Для открытой популяции: прирост = (рождаемость + иммиграция) – (смертность + эмиграция).
5. Абсолютный прирост популяции. Этот показатель представляет собой это изменение абсолютной численности популяции за бесконечно малый промежуток времени. Абсолютное изменение численности популяции в единицу времени обозначается выражением dN/dt.
Если относительный прирост популяции (r) – величина постоянная (не зависит от численности популяции), то изменение численности популяции описывается уравнением неограниченного (экспоненциального) роста. В этом случае численность популяции в данный момент времени зависит, в первую очередь, от величины r.
Однако в реальных сообществах всегда существуют лимитирующие факторы, ограничивающие численность популяций. Максимально возможная численность популяции в данных условиях называется ёмкость экологической ниши и обозначается символом К. В этом случае изменение численности популяции описывается логистическим уравнением, которому соответствует кривая ограниченного роста.
Популяции видов, у которых рождаемость и смертность в значительной мере зависят от действия внешних факторов, быстро изменяют свою численность. Периодические изменения численности популяций называются популяционными волнами. В некоторых случаях численность изменяется в тысячи и миллионы раз. Эти популяции редко достигают численности К и существуют за счет высокого значения r. Такой способ воспроизведения популяций называется r–стратегия.
r–Стратеги (эксплеренты) характеризуются низкой конкурентоспособностью, высокой плодовитостью, отсутствием заботы о потомстве, быстрым развитием и короткой продолжительностью жизни. r–Стратегов образно называют «шакалами», поскольку они способны за короткое время завоевывать освободившееся экологическое пространство.
Популяции видов, у которых рождаемость и смертность в значительной мере зависят от их плотности (то есть от характеристики самой популяции), в меньшей степени зависят от действия внешних факторов. Они поддерживают численность, близкую к величине К, поэтому способ воспроизведения таких популяций называется К–стратегия.
К–Стратеги (виоленты) характеризуются высокой конкурентоспособностью, низкой плодовитостью, заботой о потомстве, длительным развитием и длительной продолжительностью жизни. К–Стратегов образно называют «львами», поскольку они способны долгое время удерживать экологическое пространство.
Кроме r–стратегии и К–стратегии выделяется еще и S–стратегия. S–Стратеги (патиенты) населяют местообитания с неблагоприятными условиями жизни для большинства организмов, в которых конкуренция практически отсутствует. Поэтому S–стратегов образно называют «верблюдами». По низкому значению r они близки к «львам» (виолентам), а по высокому значению К – к «шакалам» (эксплерентам). По длительности развития и длительности жизни S–стратеги могут быть сходными и с r–стратегами, и с К–стратегами.
Биологическая структура популяции
В понятие биологической структуры популяции включается ее половая, возрастная (онтогенетическая) и этологическая структура.
Половая структура популяции определяется соотношением полов, которая изменяется с возрастом:
– первичное соотношение полов – при образовании зигот;
– вторичное соотношение полов – у новорожденных (независимо от способа рождения);
– третичное соотношение полов – к моменту наступления половой зрелости.
Таким образом, половая структура популяции оказывается тесно связанной с ее возрастной структурой.
Возрастная структура популяции зависит от способа размножения, от особенностей жизненного цикла и типа онтогенеза. Иногда принадлежность к возрастной группе определяется астрономическим возрастом особи (сеголетки, двухлетки и т.д.). Чаще выделяют следующие возрастные группы:
1. Новорожденные особи, или особи нулевого возраста (отложенные яйца, вылупившиеся личинки, споры или семена растений, проростки растений).
2. Особи на раннем прегенеративном (ювенильном) этапе онтогенеза; они значительно отличаются от взрослых особей.
3. Особи на позднем прегенеративном (имматурном, виргинильном) этапе онтогенеза; они сходны с взрослыми особями, но еще не могут размножаться.
4. Особи на генеративном этапе онтогенеза, которые активно размножаются.
5. Особи на постгенеративном (сенильном) этапе онтогенеза, которые уже не могут размножаться.
В конкретных случаях эта классификация уточняется. Иногда выделяют неполночленные популяции, в которых отсутствует та, или иная возрастная группа. Например, многие беспозвоночные и однолетние растения зимуют на стадии яйца или семян.
Взаимосвязь половой и возрастной структуры отражается в виде половозрастных пирамид. Пример половозрастной пирамиды для нормальной полночленной популяции показан на рисунке.
Наличие сложной половозрастной структуры обусловливает внутрипопуляционную дифференциацию экологических ниш, особенно, у животных с полным метаморфозом. Экологическая ниша всей популяции распадается на субниши, которые занимают особи, принадлежащие к разным половозрастным группам. При дифференциации ниш происходит снижение уровня конкуренции, в первую очередь, топической и трофической.
Приведем примеры дифференциации внутривидовых ниш у животных:
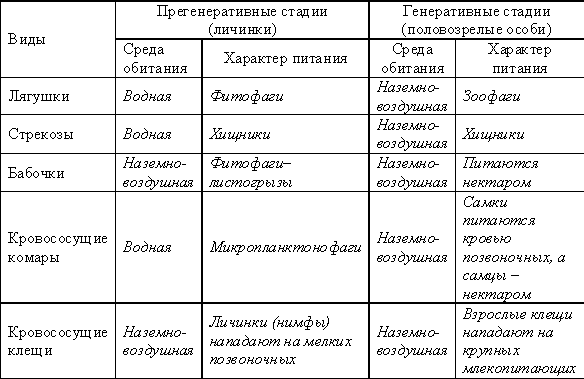
Этологическая (поведенческая) структура популяций у животных обусловлена наличием внутрипопуляционных группировок, члены которых непосредственно взаимодействуют между собой. Такие группировки называют демы (семьи), соседства, субпопуляции, парцеллы, локусы.
Существует множество уровней агрегации особей: одиночные особи, семьи, колонии, стада, стаи.
Одиночные особи могут существовать ограниченное время; по крайней мере, в период размножения неизбежно существование брачных пар и появление самок с потомством. Поэтому одиночный образ жизни периодически становится семейным. В состав семьи входят особи, связанные ближайшей степенью родства: родители и их непосредственные потомки. Существуют семьи с участием только одного из родителей (обычно, матери), двух родителей (моногамные), нескольких родителей (полигамные), с участием потомков из нескольких приплодов.
На уровне одиночных особей и семей проявляется наличие индивидуальных участков и охраняемых территорий. Индивидуальный участок – это часть территории (акватории), на которой особь проводит значительную часть времени. Охраняемая территория отличается от индивидуального участка тем, что ее владелец активно защищает свои владения.
Колонии – групповые поселения оседлых животных. Выделяют постоянные колонии (например, у грызунов) и временные (например, у птиц в период размножения). К колониям формально относятся поселения пчел, муравьев, термитов (на самом деле, это семьи).
Стада – более или менее постоянные объединения подвижных животных. Немногочисленные стада с нарушенным соотношением полов фактически являются семьями; например, прайды львов состоят из 1-3 самцов, нескольких самок и детенышей. Для стад характерно наличие строгой иерархии – порядка соподчинения особей.
Стаи – временные объединения животных, например, на определенных стадиях развития (мальки рыб) или в определенные сезоны (птицы, псовые). Стаи обеспечивают определенные функции вида: защиту от врагов, добычу пищи, миграции. Уровень соподчинения в стаях меньше, чем в стадах, и может вообще отсутствовать.
В любом случае, внутрипопуляционная группировка обладает признаками, отсутствующими у отдельных особей. В группе облегчается добывание пищи, происходит обмен пищей, повышается защищенность от неблагоприятных факторов. Эффект группы выражается в том, что происходит оптимизация физиологических процессов: например, снижается потребление кислорода, уменьшается частота сердечных сокращений, частота дыхания, повышается вероятность успешного размножения. Однако при чрезмерном увеличении численности группы возрастает внутривидовая конкуренция, и перечисленные положительные эффекты сменяются прямо противоположными.