Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Методы изучения и диагностики внимания
|
|
Экспериментальное изучение физиологических коррелятов и механизмов внимания осуществляется на разных уровнях, начиная от нервной клетки и кончая биоэлектрической активностью мозга в целом. Каждый из этих уровней исследования формирует свои представления о физиологических основах внимания.
Нейроны новизны. Наиболее интересные факты, иллюстрирующие функции нейронов в механизмах внимания, связаны с обеспечением ориентировочной реакции. Еще в 60-е гг. Г. Джаспер во время нейрохирургических операций выделил в таламусе человека особые нейроны — " детекторы" новизны, или внимания, которые реагировали на первые предъявления стимулов.
Позднее в нейронных сетях были выделены нервные клетки, получившие название нейронов новизны и тождества (Е.Н. Соколов, 1995). Нейроны новизны позволяют выделять новые сигналы. Они отличаются от других характерной особенностью: их фоновая импульсация возрастает при действии новых стимулов разной модальности. С помощью множественных связей эти нейроны соединены с детекторами отдельных зон коры головного мозга, которые образуют на нейронах новизны пластичные возбуждающие синапсы. Таким образом, при действии новых стимулов импульсная активность нейронов новизны возрастает. По мере повторения стимула и в зависимости от силы возбуждения ответ нейрона новизны избирательно подавляется, так, что дополнительная активация в нем исчезает и сохраняется лишь фоновая активность.
Нейрон тождества также обладает фоновой активностью. К этим нейронам через пластичные синапсы поступают импульсы от детекторов разных модальностей. Но в отличие от нейронов новизны, в нейронах тождества связь с детекторами осуществляется через тормозные синапсы. При действии нового раздражителя фоновая активность в нейронах тождества подавляется, а при действии привычных раздражителей, напротив, активизируется.
Итак, новый стимул возбуждает нейроны новизны и тормозит нейроны тождества, таким образом новый раздражитель стимулирует активирующую систему мозга и подавляет синхронизирующую (тормозную) систему. Привычный стимул действует прямо противоположным образом — усиливая работу тормозной системы, не влияет на активирующую.
Особенности импульсной активности нейронов человека при выполнении психологических проб, требующих мобилизации произвольного внимания, описаны в работах Н.П. Бехтеревой и ее сотрудников. При этом в передних отделах таламуса и ряде других структур ближайшей подкорки были зафиксированы стремительные возникающие вспышки импульсной активности, по частоте в 2-3 раза превышающие уровень фона. Характерно, что описанные изменения в импульсной активности нейронов сохранялись на протяжении выполнения всего теста, и только по его завершении уровень активности этих нейронов возвращался к исходному.
В целом в этих исследованиях установлено, что различные формы познавательной деятельности человека, сопровождающиеся напряжением произвольного внимания, характеризуются определенным типом нейрональной активности, четко сопоставимым с динамикой произвольного внимания.
Электроэнцефалографические корреляты внимания. Хорошо известно, что при предъявлении стимула в энцефалограмме наблюдается подавление (блокада) альфа-ритма, и на смену ему приходит реакция активации. Однако этим не исчерпываются изменения электрической активности мозга в ситуации внимания.
Исследование суммарной электрической активности при мобилизации интеллектуального внимания выявило закономерные изменения в характере совместной деятельности разных зон коры. При оценке степени дистантной синхронизации биопотенциалов было установлено, что в передних зонах левого полушария существенно по сравнению с фоном увеличивается уровень пространственной синхронизации. Сходные результаты дает использование и другого показателя, извлекаемого из энцефалограммы, — когерентности (см. тему 2 п. 2.1.1). В ситуации ожидания стимула независимо от его модальности наблюдается рост когерентности в полосе альфа-ритма, причем преимущественно в передних (премоторных) зонах коры. Высокие показатели дистантной синхронизации и когерентности говорят о том, насколько тесно взаимодействуют зоны коры, в первую очередь передних отделов левого полушария, в обеспечении произвольного внимания.
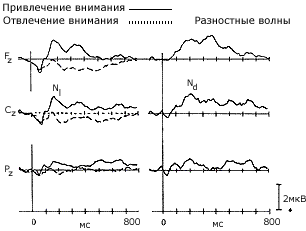
| Слуховые вызванные потенциалы, отражающие привлечение селективного внимания к одному из каналов в ситуации различения звуковых сигналов (700 или 300 Гц) (по H. Hansen & S. Hillyard, 1982). Высоко- и низкочастотные тоны предъявлялись в случайном порядке (приблизительно три раза в сек.). Испытуемые каждый раз обращали внимание только на один канал, пытаясь выделить сигнальные стимулы, имевшие большую длительность ВП в канале, к которому было привлечено внимание, имели выраженную негативную волну. Эта волна отчетливо выступает при вычитании ответа на сигнальный стимул из ответа на несигнальный - на рис. справа. |
Изучение внимания с помощью ВП. Первые исследования внимания методом ВП использовали простые поведенческие модели, например, счет стимулов. При этом было установлено, что привлечение внимания испытуемых к стимулу сопровождается увеличением амплитуды компонентов ВП и сокращением их латентности. Напротив, отвлечение внимания от стимула сопровождается снижением амплитуды ВП и увеличением латентности. Однако оставалось неясным, чем обусловлены эти изменения параметров ВП: изменением общего уровня активации, поддержанием бдительности или механизмами избирательного внимания. Для разведения указанных процессов необходимо было построить эксперимент таким образом, чтобы его организация позволяла вычленить эффект мобилизации селективного внимания " в чистом" виде.
В качестве такой модели можно привести эксперименты С. Хильярда, которые получили в 70-е гг. широкую известность. При предъявлении звуковых стимулов через наушники в левое и правое ухо испытуемому предлагается мысленно реагировать (считать) редко встречающиеся (" целевые") стимулы, поступающие по одному из каналов (только в правое или левое ухо). В результате получают вызванные потенциалы в ответ на 4 варианта стимулов: часто встречающиеся в релевантном (контролируемом) и иррелевантном (игнорируемом) каналах и редко встречающиеся (целевые) в том и другом каналах. В этом случае появляется возможность сравнивать эффекты канала и стимула, которые являются объектом внимания. В экспериментах такого типа, как правило, применяются очень короткие интервалы между стимулами (немногим более или менее одной секунды), в результате усиливается напряженность и устойчивость избирательного внимания испытуемого к быстро чередующимся стимулам разной информационной значимости.
 Было установлено, что привлечение внимания к одному из каналов ведет к увеличению амплитуды первой отрицательной волны с латентным периодом около 150 мс, обозначаемой как компонент N1. Целевые стимулы сопровождались появлением в составе ВП позднего положительного колебания Р3 с латентным периодом около 300 мс. Высказывалось предположение, что негативная волна N1 отражает " установку" на стимул, определяющую направленность произвольного внимания, а компонент Р3 — " установку на ответ", связанную с выбором варианта ответа. В дальнейшем компонент Р3 (чаще определяемый как Р300) явился предметом множества исследований (см. тему 10).
Было установлено, что привлечение внимания к одному из каналов ведет к увеличению амплитуды первой отрицательной волны с латентным периодом около 150 мс, обозначаемой как компонент N1. Целевые стимулы сопровождались появлением в составе ВП позднего положительного колебания Р3 с латентным периодом около 300 мс. Высказывалось предположение, что негативная волна N1 отражает " установку" на стимул, определяющую направленность произвольного внимания, а компонент Р3 — " установку на ответ", связанную с выбором варианта ответа. В дальнейшем компонент Р3 (чаще определяемый как Р300) явился предметом множества исследований (см. тему 10).
В более поздних исследованиях с помощью специального приема вычитания потенциалов, регистрируемых в ответ на сигнальные и стандартные стимулы, обнаружилось, что первая отрицательная волна N1 представляет собой неоднородный корковый феномен сложной структуры, в котором можно выделить особое отрицательное колебание, так называемую — " негативность, отражающую обработку информации". Это колебание с латентным периодом около 150 мс и длительностью не менее 500 мс регистрируется при несовпадении редко предъявляемого целевого стимула со " следом внимания", образуемым в ассоциативной слуховой зоне и лобной области при частом повторении и воспроизведении стандартного стимула. При этом, чем меньше разница между этими стимулами, тем больше латентный период и тем длительнее отрицательное колебание, развивающееся в ответ на целевой, нестандартный стимул.
Кроме этого, описано еще одно отрицательное колебание, в ряде случаев сопровождающее ситуацию сравнения стимулов. Этот компонент, обозначаемый как " негативность рассогласования ", возникает в слуховой коре с латентным периодом 70-100 мс и отражает автоматический процесс сравнения физических признаков звукового стимула со следом стандартного стимула, хранящемся в течение 5-10 сек в сенсорной памяти. При отклонении физических свойств стимула от следа многократно предъявляемого стандартного стимула развивается " негативность рассогласования".
Предполагается, что в образовании волны N1 могут участвовать оба компонента (" негативность, связанная с обработкой информации" и " негативность рассогласования"). Причем первый из этих компонентов связан с предсознательной, непроизвольной оценкой признаков необычного звукового стимула, осуществляемой путем сравнения их с нервной моделью часто повторяющегося стимула, а второй компонент отражает процессы обработки сенсорной информации на сознательном уровне, а именно: произвольного внимания, фокусирования субъектом сознания на определенных критических признаках стимула и сравнения его со " следом внимания", хранящемся в рабочей памяти.
Таким образом, с помощью метода ВП было показано, что на целевые звуковые стимулы (в ситуации выбора стимула и канала) возникает два типа компонентов, один из которых отражает процессы сенсорной памяти, другой — селективного внимания.
Временные характеристики внимания. С помощью метода ВП можно оценить динамику развития процессов внимания в реальном времени. Вопрос заключается в следующем, на каком этапе обработки информации включаются процессы внимания? Поскольку начало первой негативной волны, возникающей в ответ на сигнальные стимулы, в основном приурочено к 50 мс от момента предъявления стимула, пятидесятимиллисекундная граница довольно долго рассматривалась как временной рубеж, после которого развертываются процессы селективного внимания.
Более детальные исследования, однако, показали, что в слуховой и, по-видимому, соматосенсорной системах произвольная регуляция процессов обработки поступающей информации включается не позже, чем через 20-30 сек. после предъявления стимула. Эффекты внимания в зрительной системе обнаруживают себя позднее, начиная с 60 мс. Не исключено, что и эти временные границы по мере совершенствования методов изучения будут изменены. Суть, однако, в том, что хронометрия переработки информации и включения внимания как одного из главных регуляторов этого процесса с такой точностью может быть изучена только в психофизиологических экспериментах.