Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Г. Строение 3'- и 5'-концов цепей ДНК. Теломерная ДНК
|
|
На каждом конце хромосомы присутствует специфическая нуклеотидная последовательность. Она представлена многочисленными повторами (сотни или даже тысячи раз) олигонуклеотидов
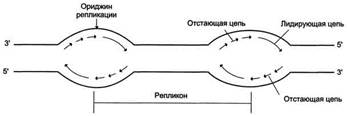
Рис. 4-17. Образование двух репликативных вилок, перемещающихся в противоположных направлениях от ориджина.
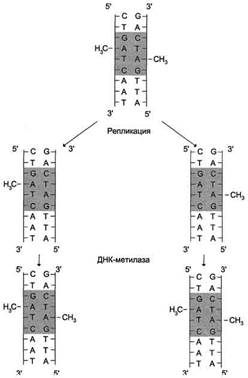
Рис. 4-18. Метилирование остатков аденина в последовательности -GATC-. В течение нескольких минут после репликации, пока не произошло метилирование, новая цепь ДНК отличается от матричной цепи.
-GGGTTA-, называемых теломерной последовательностью, или просто теломерной ДНК. Наличие теломер необходимо для завершения репликации концевых информативных последовательностей хромосом, т.е. для сохранения генетической информации.
После завершения репликации хромосомы 5'-концы дочерних цепей ДНК недостроены, так как после удаления праймеров эти фрагменты оказываются недореплицированными. Это происходит потому, что ДНК-полимераза β, отвечающая за заполнение бреши, образованной после удаления праймера, не может вести синтез цепи ДНК от 3'- к 5'-концу (рис. 4-19, А). Таким образом, в ходе каждого цикла репликации 5'-концы синтезированных цепей укорачиваются. Но такие потери не представляют опасности для генетической информации хромосом, потому что укорочение ДНК идёт за счёт теломер. Во время следующего цикла репликации
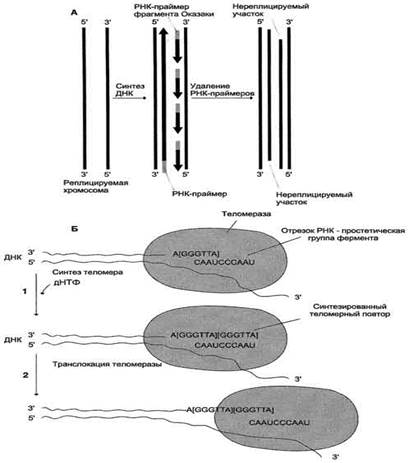
Рис. 4-19. Синтез теломерной ДНК. А - на рисунке показано укорочение вновь синтезированных цепей ДНК после удаления праймеров; Б - в состав теломеразы входит короткая молекула РНК, содержащая в активном центре последовательность нук-леотидов, комплементарную теломерному повтору; 1 - фермент прикрепляется за счёт взаимодействия РНК с существующей теломерой и добавляет последовательно по одному нуклеотиду фрагмент -GGGTTA-. Матрицей служит простетическая группа теломеразы - фрагмент РНК; 2 - фермент перемещается по нити ДНК таким образом, что РНК-матрица в составе теломеразы постоянно комплементарно связана с концом вновь синтезированного теломерного повтора. Заново синтезированная тело-мерная ДНК служит матрицей для удлинения второй цепи ДНК, но уже в ходе следующего цикла клеточного деления. Теломер-ный повтор на рисунке взят в квадратные скобки -[GGGTTA]-.
5'-концы цепей ДНК опять остаются недостроенными. Таким образом, с каждым клеточным делением ДНК хромосом будут последовательно укорачиваться. Укорочение теломер в большинстве клеток по мере их старения - важный фактор, определяющий продолжительность жизни организма.
Однако в эмбриональных и других быстро-делящихся клетках потери концов хромосом недопустимы, потому что укорочение ДНК будет происходить очень быстро. В эукариотических клетках имеется фермент теломераза (нуклео-тидилтрансфераза), обеспечивающий восстановление недореплицированных 5'-концов. К особенностям этого фермента относят присутствие в качестве простетической группы РНК. Фрагмент РНК в активном центре теломеразы служит матрицей при синтезе теломер-ных повторов хромосом.
С помощью РНК фермент комплементарно прикрепляется к 3'-концу недостроенной дочерней цепи ДНК. Теломераза по принципу комплементарности последовательно удлиняет 3'-конец цепи ДНК на один гексануклеотид -GGGTTA-. Синтез всегда идёт от 5'- к 3'-концу. Затем теломераза смещается по цепи ДНК на один теломер и начинает синтез нового фрагмента -GGGTTA- (рис. 4-19, Б).
В большинстве соматических клеток теломераза неактивна, так как соматическая клетка имеет длину теломерной ДНК, достаточную для времени жизни клетки и её потомства. Однако небольшую активность теломеразы обнаруживают в клетках с высокой скоростью обновления, таких как лимфоциты, стволовые клетки костного мозга, клетки эпителия, эпидермиса кожи и др.