Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Нейронные цепи
|
|
Нервная система человека может быть представлена в виде нервной сети, т.е. системы нейронных цепочек, передающих возбуждающие и тормозные сигналы. Нейронные сети построены из трех главных компонентов: входных волокон, вставочных нейронов и эфферентных нейронов. Самыми простыми и элементарными нейронными цепями являются локальные сети, или микросети (рис. 69). Нередко определенный тип микросети повторяется по всему слою нервной структуры, например коры больших полушарий, и выступает в качестве модуля для особого способа обработки информации.
Локальные сети имеются в разных отделах мозга. Они служат: 1) для усиления слабых сигналов; 2) уменьшения и фильтрации слишком интенсивной активности; 3) выделения контрастов; 4) поддержания ритмов или сохранения рабочего состояния нейронов путем регулировки их входов. Микросети могут оказывать на нейроны-мишени возбуждающее или тормозящее действие.
Локальные сети можно сравнить с интегральными микросхемами в электронике, т.е. стандартными элементами, которые выполняют наиболее часто повторяющиеся операции и могут быть включены в схемы самых разнообразных электронных приборов.
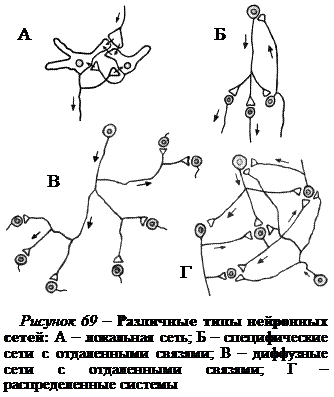 Одну из разновидностей локальных сетей составляют, как правило, нейроны с короткими аксонами (рис. 69, А). Поэтому задачи и сферы влияния таких нейронов весьма ограничены. Вторую разновидность локальной сети образуют нейроны, достаточно удаленные друг от друга, но принадлежащие к одной нервной области. Основными функциями этих сетей является распространение активности за пределы отдельного модуля или же обеспечение антагонистических взаимодействий между соседними модулями в пределах данной нервной области.
Одну из разновидностей локальных сетей составляют, как правило, нейроны с короткими аксонами (рис. 69, А). Поэтому задачи и сферы влияния таких нейронов весьма ограничены. Вторую разновидность локальной сети образуют нейроны, достаточно удаленные друг от друга, но принадлежащие к одной нервной области. Основными функциями этих сетей является распространение активности за пределы отдельного модуля или же обеспечение антагонистических взаимодействий между соседними модулями в пределах данной нервной области.
Более сложными являются сети с отдаленными связями, соединяющие две или несколько областей нервной системы с локальными сетями. Сети с отдаленными связями могут быть как специфическими (рис. 69, Б), так и диффузными (рис. 69, В). Специфическое последовательное соединение нескольких областей выполняет функцию передачи информации с периферии в центральную нервную систему (например, проводниковые отделы анализаторов) или с центральных отделов на периферию (например, моторная система). В таких случаях обычно сети с отдаленными связями называют восходящими и нисходящими путями, или системами. Нервные структуры, входящие в восходящие пути, объединяются по принципу восходящей иерархии, а образующие нисходящие пути – по принципу нисходящей иерархии.
Самый высокий уровень организации – это система соединений между целым рядом областей, управляющих каким-то поведением, в котором участвует весь организм. Такие сети носят название распределенных систем (рис. 69, Г). Они могут находиться в разных отделах мозга и могут быть связаны гормональными воздействиями или длинными нервными путями. Распределенные системы участвуют в осуществлении высших функций двигательных и сенсорных систем, а также многих других центральных систем, обеспечивающих сложные поведенческие акты, абстрактное мышление, речь и другие психофизиологические процессы.
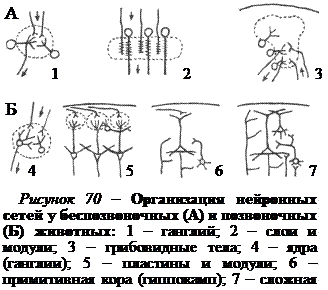 В процессе эволюции происходило усложнение нейронных сетей. У беспозвоночных животных со слабо интегрированной нервной системой нейронные сети обычно организованы либо в форме ганглиев, либо в форме пластинок (рис. 70, А). Ганглии являются структурой с концентрированным расположением синаптических контактов между входными и выходными элементами, а пластинки – структурой с двухслойной организацией таких контактов.
В процессе эволюции происходило усложнение нейронных сетей. У беспозвоночных животных со слабо интегрированной нервной системой нейронные сети обычно организованы либо в форме ганглиев, либо в форме пластинок (рис. 70, А). Ганглии являются структурой с концентрированным расположением синаптических контактов между входными и выходными элементами, а пластинки – структурой с двухслойной организацией таких контактов.
У высших беспозвоночных животных интеграция сигналов на более высоком уровне происходит в нервных центрах, примером чему являются грибовидные тела мозга у насекомых. Грибовидные тела спрятаны в глубине мозга, а не на его поверхности, где они могли бы разрастаться в ширину.
У позвоночных животных и у человека часть нейронных сетей группируются в ганглии. Центры, расположенные в глубине мозга, увеличиваются за счет образования изгибов, подобно грибовидным телам насекомых. Однако принципиально новая особенность высших позвоночных и
человека – это группировка огромного количества нейронов в слои, лежащие на поверхности мозга, т.е. образование коры (рис. 70, Б).
Кора расположена таким образом, что нейроны всех ее слоев доступны для любых входных сигналов. Совместно с локальными сетями, образованными ответвлениями отростков нейронов и вставочными нейронами, кора обладает огромными возможностями для интеграции, хранения и комбинирования информации. В каждом участке, или поле коры, многократно повторяются однотипные модули (локальные сети), благодаря которым данное поле способно осуществлять специфические операции с участием определенных входных и выходных связей (зрительное поле, слуховое поле). При переходе к соседнему полю коры все эти три элемента, т.е. локальные сети, входы и выходы, слегка изменяются. Изменяются и функциональные свойства. Таким образом, каждое из полей коры является участком, приспособленным для выполнения определенных функций в той распределенной системе, в состав которой оно входит.
5.11. Основные законы функционирования нервных сетей
5.11.1. Дивергенция и конвергенция нервных путей
Во всех изученных нервных сетях обнаружены дивергенция и конвергенция путей. Дивергенцией называют способность нейрона устанавливать многочисленные синаптические связи со многими другими нервными клетками (рис. 71, А). Так, например аксон чувствительного нейрона входит в задние рога спинного мозга в составе задних корешков и в спинном мозге разветвляется на множество веточек (коллатералей), образующих синапсы на многих вставочных нейронах и мотонейронах. Благодаря процессу дивергенции одна и та же нервная клетка может участвовать в различных нервных реакциях и контролировать большое число других нейронов. Такое расширение сферы действия и распространение сигнала в нервных сетях называется иррадиацией. Иррадиировать может как возбуждение так и торможение.
Схождение многих нервных путей к одному и тому же нейрону носит название конвергенции (рис. 71, Б). Например, на каждом мотонейроне спинного мозга образуют синапсы тысячи отростков чувствительных, а также возбуждающих и тормозных вставочных нейронов из разных отделов ЦНС. Вследствие конвергенции многих нервных путей к одному нейрону этот нейрон осуществляет интеграцию одновременно поступающих по разным путям возбуждающих и тормозных сигналов. Если в результате алгебраического сложения ВПСП и ТПСП, возникающих на мембране нейрона, возбуждение будет преобладать, то нейрон возбудится и пошлет нервный импульс ко второй клетке. Если будет преобладать достаточной величины ТПСП, то нейрон затормозится. Такое сложение постсинаптических потенциалов называют пространственной, или одновременной суммацией.
 Афферентных нейронов в нервной системе примерно в 5 раз больше, чем эфферентных. В связи с этим многие афферентные импульсы поступают к одним и тем же вставочным и эфферентным нейронам, которые являются для импульсов общими конечными путями к рабочим органам.
Афферентных нейронов в нервной системе примерно в 5 раз больше, чем эфферентных. В связи с этим многие афферентные импульсы поступают к одним и тем же вставочным и эфферентным нейронам, которые являются для импульсов общими конечными путями к рабочим органам.
Впервые закономерности общих конечных путей были изучены в начале 20-го века английским физиологом Ч. Шеррингтоном. Морфологической основой общих конечных путей является конвергенция нервных волокон. Благодаря общим конечным путям одна и та же ответная рефлекторная реакция определенной группы мотонейронов может быть получена при раздражении различных нервных структур. Например, мотонейроны, иннервирующие мышцы глотки, участвуют в рефлексах глотания, кашля, сосания, дыхания, образуя общий конечный путь для различных рефлекторных дуг.
Рефлексы, дуги которых имеют общий конечный путь, подразделяются на союзные и антагонистические. Встречаясь на общих конечных путях, союзные рефлексы взаимно усиливают друг друга, а антагонистические – тормозят друг друга, как бы конкурируя за захват общего конечного пути. Преобладание на конечных путях той или иной, в том числе и поведенческой, рефлекторной реакции обусловлено ее значением для жизнедеятельности организма в данный момент.