Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Рзповсюдження
|
|
В першій половині ХХ сторіччя вважалося, що передача нервового сигналу за допомогою хімічного синапсу притаманна периферійним нервово-м’язовим контактам хребетних (автономна мускулатура та моторні контакти), але, в основному через брак дослідних даних, синапси в Центральній нервовій системі (ЦНС) вважались такими, що використовують безпосередньо електричну передачу. Але, після відпрацювання методик внутрішньоклітинного запису електричних потенціалів в 1950-х роках, було показано, що з’єднання нейронів хімічними синапсами є правилом для ЦНС хребетних.
Надалі деякі синапси в нервовій системі ракоподібних були ідентифіковані як електричні. На теперішній час виявлена невелика кількість електричних синапсів в мозку ссавців, і велика їх доля в ЦНС нижчих хребетних, особливо риб. У безхребетних електричні синапси досить звичайні в нервовій системі аннелід та ракоподібних. Електричні синапси звичайно розташовані на таких нервових шляхах, де потрібне додержання точно визначеного часу проходження нервового імпульсу – з огляду на те, що електричний синапс не спричинює помітної в досліді затримки сигналу.
Будова
На відміну від хімічного синапсу, синаптична щілина в електричному синапсі є надзвичайно вузькою (близько 3.5 нанометри). Через синаптичну щілину даного типу синапсів проходять просторво впорядковані гідрофільні протеїнові тунелі, кожний приблизно 5 нанометрів завширшки, котрі перфорують пре- та постсинаптичну мембрану і називаються коннексонами. У первинноротих організмів (нематоди, молюски, членистоногі) коннексони сформовані протеїнами специфічної структури, що називаються паннексинами або іннексинами; у вторинноротих (голкошкірі, асцидії, хребетні) коннексони побудовані з протеїнів іншого типу – коннексинів, котрі кодуються іншою групою генів.
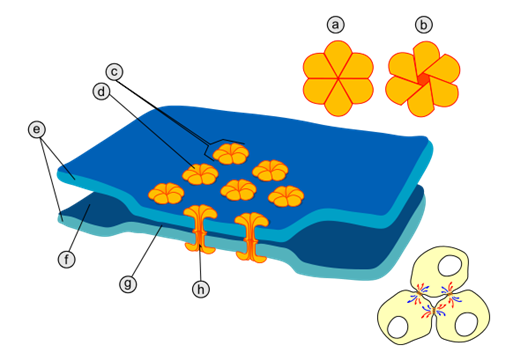 |
Рис.2 Будова електричного синапсу: а - коннексон у закритому стані; b - коннексон у відкритому стані; с - коннексон, вбудований в мембрану; d - мономер коннексину; е - плазматична мембрана; f - міжклітинний простір; g - проміжок у 2-4 нанометри в електричному синапсі; h - гідрофільний канал коннексону.
Ссавці в цьому плані є менш диференційовані вториннороті, і в їхньому організмі поряд з коннексинами виробляються також і паннексини, але досі у хребетних не виявлено жодного електричного синапсу, де міжклітинні канали були б сформовані паннексинами.
Міжмембранні тунелі, сформовані коннексинами (або паннексинами) забезпечують рідинний взаємозв’язок між двома нейронами – пре- та постсинаптичним - та уможливлюють прохід через них іонів та малих молекул, в тому числі штучно введених в клітину флуоресцентних барвників. Прохід вказаних барвників через електричний синапс може бути зареєстрований навіть за допомогою світлового мікроскопу.
Електричні синапси дозволяють здійснення електричної провідності в обох напрямках (на відміну від хімічних); втім, останнім часом у деяких ракоподібних було відкрито спрямовуючі електричні синапси, тобто такі, що дозволяють проходження нервового сигналу тільки в одному напрямку.
3.3. Функції
Описані властивості електричних синапсів визначають їхні функції в нервовій системі різних організмів.
Завдяки прямому переходу іонів через такий синапс нервовий сигнал передається ними практично без затримки. Це дозволяє ракоподібним, у яких такі синапси поєднують нейрони, що відповідають за рухову активність, мінімізувати час між появою небезпеки та моторною реакцію на неї – що часто є критичним при втечі від хижака.
Більш загальна функція електричних синапсів, основана на проведенні ними сигналу в обох напрямках, полягає в синхонізації активності нейронних популяцій. Наприклад, нейрони стволу головного мозку, що генерують ритмічні електричні імпульси, які забезпечують дихання, вочевидь мають синхронизувати цю активність. Така синхронізація досягається завдяки наявності в їхній популяції електричних синапсів, які миттєво збуджують клітини, що „відстали” на якому-небудь такті збудження від інших. Популяції нейронів, де наявні електричні синапси, які забезпечують синхронізацію збудження, виявлені також в корі, таламусі, мозочку та інших частинах мозку.
Той факт, що розмір пори коннексонів електричних синапсів дозволяє перехід з клітини в клітину молекул АТФ та вторинних месенджерів, також відіграє важливу роль в забезпеченні синхронізації збудження та метаболізму нейронів - останнє особливо важливо для клітин нейроглії, поєднання котрих електричними синапсами було відкрите нещодавно.
4. Змішаний синапс
Змішані синапси - синапси, в котрих одночасно наявна електрична та хімічна провідність нервового сигналу; таким чином, вони сполучають функції та будову хімічного та електричного синапсу.
Змішані синапси виявлені майже винятково в нервовій системі риб.
Функції та роль змішаних синапсів наразі вивчені слабо.
Література
1. Anat Rec B New Anat. 2005 Porocytosis: a transient pore array secretes the neurotransmitter packet.
2. Brain Res. 1997 Dynamic responses of presynaptic terminal membrane pools following KCl and sucrose stimulation.
3. Костюк П.Г. та ін. Біофізика // Київський університет / 2008, – С 567.
4. Мотавкин П.А. Курс лекций по гистологии.. — «Медицина ДВ», 2007. — С. 137.
5. Савельев А. В.. Методология синаптической самоорганизации и проблема дистальных синапсов нейронов // Журнал проблем эволюции открытых систем. — Казахстан, Алматы: 2006. — Т. 8. — № 2. — С. 96-104.
6. Экклз Д.К. Физиология синапсов. М.: Мир, 1966, 397 с.