Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Физиология дыхания
|
|
Дыхание включает следующие процессы: 1) легочное дыхание, 2) транспорт газов кровью, 3) обмен газов между кровью и тканями, 4) окисление органических веществ в клетках (тканевое дыхание). Физиология рассматривает три первых процесса, тканевое дыхание изучается в курсе биохимии.
Легочное дыхание. Механизмы вдоха и выдоха. Легочное дыхание включает обмен воздуха между окружающей средой и легкими (внешнее дыхание) и обмен газов между альвеолярным воздухом и кровью. Атмосферный воздух поступает в легкие через воздухоносные пути во время вдоха, при выдохе воздух с повышенным содержанием углекислого газа удаляется тем же путем в окружающую среду. В легких происходит диффузия кислорода в кровь и диффузия углекислого газа из крови в альвеолярный воздух.
Механизм вдоха. Акт вдоха обеспечивается сокращением наружных межреберных мышц и диафрагмы. Межреберные мышцы приподнимают ребра, несколько поворачивают их вокруг оси и отводят в стороны, а грудину — вперед. В результате объем грудной полости увеличивается в переднезаднем и боковых направлениях. Одновременно сокращается диафрагма, что приводит к снижению ее уровня на 3—4 см, увеличению размера грудной полости в вертикальном направлении и ее объема почти на 1000 мл (рис. 63). Опускаясь, диафрагма давит на органы брюшной полости, что влечет за собой выпячивание передней брюшной стенки. При усиленном глубоком вдохе сокращаются и некоторые другие мышцы брюшного пресса и груди.
Легкие постоянно находятся в растянутом состоянии. Это происходит потому, что рост грудной клетки после рождения и в процессе развития ребенка опережает рост легких. Кроме того, атмосферный воздух производит одностороннее (изнутри) давление на легкие через воздухоносные пути. Грудная клетка неподатлива к передаче давления воздуха снаружи на легкие, поэтому атмосферный воздух, растягивая легкие, прижимает их к плевре и грудной стенке. Вследствие этого по форме плевральная полость представляет собой узкую, герметически закрытую щель между ее висцеральным и париетальным листками. Однако сама легочная ткань, обладая большой эластичностью, противодействует растяжению. Эта так называемая эластическая тяга легких уменьшает величину давления воздуха на легкие и плевру.
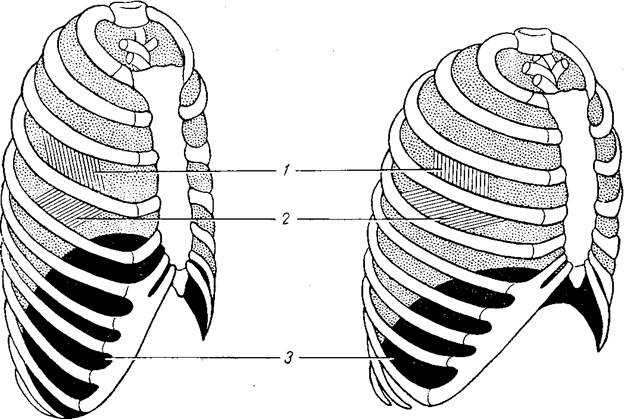
Рис. 63. Положение грудной клетки и диафрагмы при выдохе (слева) и при вдохе (справа) (схема).
1 — наружные межреберные мышцы; 2 — внутренние межреберные мышцы; 3 — диафрагма.
Давление в плевральной полости ниже, чем атмосферное, на 4—9 мм рт. ст. Поэтому плевральное давление называют отрицательным, условно принимая атмосферное давление (760 мм рт. ст.) ра нулевое. Чем сильнее растягиваются легкие, тем выше становится их эластическая тяга и тем ниже падает давление в плевральной полости. Во время вдоха оно равно 9 мм рт. ст., во время выдоха 4 мм рт. ст., а при глубоком вдохе давление может падать до 3 мм рт. ст.
Отрицательное давление в плевральной полости обеспечивается также деятельностью мезотелиальных клеток, выстилающих плевральную полость, так как они обладают способностью поглощать воздух.
Поступление воздуха в легкие является пассивным процессом и обусловлено разностью давления его в легких и окружающей среде. При вдохе объем грудной полости увеличивается, давление в плевральной полости становится более отрицательным. Вследствие этого сопротивление в легких к растяжению уменьшается, и они растягиваются. Объем воздуха в легких увеличивается и давление воздуха в легких уменьшается, становясь ниже атмосферного. Разность давления воздуха в атмосфере и в альвеолах легких является непосредственной причиной движения воздуха из окружающей среды в легкие — происходит вдох. При вдохе тяга грудной клетки, обусловленная силой сокращения дыхательных мышц, направлена в противоположную от тяги легких сторону. Это облегчает вдох. Соотношение указанных сил определяет уровень спокойного дыхания и то количество воздуха, которое поступает в легкие при спокойном вдохе. В конце вдоха эластическая тяга грудной клетки начинает противодействовать вдоху. Их направление совпадает с направлением эластической тяги легких, что препятствует продолжению вдоха.
Механизм выдоха. Акт выдоха начинается с расслабления наружных дыхательных мышц и диафрагмы. Вследствие этого под действием эластических сил легких и давления внутренних органов, а также силы тяжести грудной клетки объем ее уменьшается. При усиленном напряженном выдохе к перечисленным силам присоединяется сокращение внутренних межреберных мышц, мышц живота и некоторых других мышц, что способствует еще большему уменьшению объема грудной полости. Уменьшение объема грудной полости обусловливает повышение плеврального давления. Оно становится менее отрицательным. В результате этого и под действием эластической тяги легких объем легких уменьшается, давление воздуха в них становится выше атмосферного, и воздух удаляется в окружающую среду. Выдох заканчивается тогда, когда эластическая тяга легких уравновешивается повышающимся давлением в плевральной полости.
Пневмоторакс. При ранениях стенки грудной полости и повреждении плевры окружающий воздух поступает в плев-ральную полость в силу пониженного в ней давления. Это явление получило название пневмоторакса. Легкое при этом спадается, дыхание может прекратиться. При двустороннем ранении наступает смерть от удушья.
Искусственный пневмоторакс применяют для лечения туберкулеза легких. В этом случае прокалывают грудную стенку иглой от шприца и впускают необходимое количество воздуха в плевральную полость. Легкое до известной степени спадается, его движения ограничиваются, что способствует закрытию и заживлению патологических полостей (каверн), образовавшихся вследствие распада легочной ткани. Через некоторое время воздух из плевральной полости рассасывается и давление в ней вновь падает. В случае необходимости искусственный пневмоторакс повторяют.
Типы дыхания. В зависимости от того, какие мышцы преимущественно участвуют в акте дыхания, различают грудной, брюшной (диафрагмальный) и смешанный типы дыхания. У мужчин тип дыхания — брюшной, у женщин — грудной. Однако тип дыхания меняется в зависимости от некоторых условий и физической работы. У женщин, занимающихся физическим трудом, начинает преобладать брюшной тип дыхания. При беременности, наоборот, тип дыхания грудной, так как движение диафрагмы вниз затруднено.
Легочная вентиляция. Жизненная емкость легких. Движение воздуха в легких во время дыхания называют легочной вентиляцией. Легочная вентиляция характеризуется минутным объемом дыхания, т. е. тем количеством воздуха, которое проходит через легкие в минуту. Вычисляют минутный объем дыхания путем умножения объема воздуха, поступившего в легкие при одном вдохе, на частоту дыхательных движений в минуту.
В состоянии покоя взрослый человек вдыхает и выдыхает около 500 мл воздуха. Частота дыхания у взрослых людей 16—20 раз в минуту. Следовательно, минутный объем дыхания в среднем равен 6—8 л. У детей дыхание более частое: у новорожденного 60 раз в минуту, у ребенка 5 лет 25, у подростков 15—16 лет оно равно 16—20 раз в минуту. Дыхание резко учащается во время физической работы, спортивных упражнений и может достигнуть 40—45 раз и более в минуту.
На частоту дыхания оказывают влияние температура окружающей среды и тела, эмоции, а также многие другие факторы. У тренированных к физическим нагрузкам людей частота дыхания ниже, но минутный объем дыхания выще за счет большей глубины вдоха. Следует иметь в виду, что часть воздуха в органах дыхания находится и циркулирует в гортани, трахее, бронхах и бронхиолах, где не происходит обмен газов между воздухом и кровью. Этот-воздух называется воздухом вредного пространства. Объем указанного пространства невелик, в среднем около 140 мл. Однако следует учитывать что при каждом вдохе обновляется не весь воздух, а Меньше на эту величину. Так, при спокойном вдохе в альвеолы легких, где происходит газообмен, поступает 500—140 = 360 мл воздуха. Чем выше минутный объем воздуха за счет глубины дыхания, тем меньше относительный объем воздуха ЦредН0-го пространства. Поэтому редкое и глубокое дыхание значительно эффективнее для снабжения организма кислородом так как в этом случае вентиляция альвеол увеличивается Весьма важным является обучение рациональному Дыханию каждого человека, чему служит специальная дыхательная гимнастика.
Жизненной емкостью легких называют тот объем В()3дуХа который человек может максимально выдохнуть после максимального глубокого вдоха.
Как указывалось выше, при спокойном дыхании Человек вдыхает в среднем 500 мл воздуха. Этот объем ; воздуха называется дыхательным объемом. Если произвести усиленный вдох, то можно вдохнуть еще дополнительно около 1 Г> оо мл Этот объем воздуха носит название дополнительного объема После спокойного обычного выдоха путем усиленного выдоха еще можно выдохнуть около 1500 мл воздуха. Этот по(леднид объем называют резервным объемом воздуха. Жизнен|[ая ем_ кость легких складывается из дыхательного, дополнит<.льного и резервного объемов воздуха и равна в среднем у вз|)ослого человека 3500 мл. У людей, тренированных к физическому напряжению, жизненная емкость легких больше, чем у нетренированных. У женщин она несколько меньше, чем у Мужчин Чем выше жизненная емкость, тем лучше снабжается О), ганизм кислородом.
После самого максимального выдоха легкие не овобождаются полностью от воздуха. В них остается около 1000-1500 мл воздуха. Этот объем воздуха называется остаточным объемом. Этот воздух остается в легких и после смерти. Поэтому кусочек легких умершего человека плавает в воде. Воздуха нет в легких лишь в период внутриутробного развития. При первом вдохе новорожденного легкие навсегда заполняются воздухом. Отсутствие воздуха в легких до рождения используется судебной экспертизой для определения времени гибели ребенка. Если легкое плавает в воде, то смерть наступила после его рождения, если тонет — то во время внутриутробной жизни.
Диффузия газов в легких. В легких совершается обмен кислорода и углекислоты между воздухом и кровью. Этот обмен происходит благодаря разнице парциального давления газов в альвеолярном воздухе и в крови, протекающей в капиллярах легких.
Диффузия газов из окружающей среды в жидкость и из жидкости в воздух подчиняется законам движения газов. Если над жидкостью находится смесь газов, то каждый газ растворяется в жидкости соответственно его парциальному давлению, т. е. тому давлению, которое приходится на fero долю от общего давления смеси газов. Парциальное давление пропорционально содержанию каждого газа в смеси. Так, при атмосферном давлении, равном 760 мм рт. ст., и температуре 22° парциальное давление кислорода воздуха умеренной влажности составляет 20, 94% от 760 мм рт. ст. и равно 159 мм рт. ст. В тех же условиях парциальное давление углекислого газа составляет 0, 03% от 760 мм рт. ст., и оно равно 0, 2 мм рт. ст. Остальная часть атмосферного давления приходится на азот (557, 8 мм рт. ст.), пары воды (20 мм рт. ст.) и инертные газы: аргон, неон, гелий, криптон, ксенон и водород (6, 9 мм рт. ст.). Суммарно их парциальное давление составляет 600, 8 мм рт. ст. Таким же путем рассчитывают парциальное Давление газов в альвеолярном воздухе. Следует учитывать, что в альвеолярном воздухе присутствует большее количество паров воды, на долю которых приходится часть давления смеси газов (приблизительно 47 мм рт. ст.). В альвеолярном воз-Духе парциальное давление кислорода равно 107—110 мм рт. ст., углекислого газа — 40 мм рт. ст. и азота — 570 мм рт. ст.
Газы из области с высоким парциальным давлением переходят в область с низким парциальным давлением. Поэтому вдыхаемый (атмосферный), выдыхаемый и альвеолярный воз-Дух отличается по содержанию в нем кислорода и углекислого газа (табл. 3).
Вдыхаемый воздух содержит 20, 94% кислорода, 0, 03% углекислого газа и 79, 03% азота, т. е. богат кислородом и очень беден углекислым газом. Количество азота не изменяется при газообмене, а некоторое увеличение его процентного содержания в выдыхаемом и альвеолярном воздухе объясняется небольшим уменьшением объема выдыхаемого воздуха по сравнению с вдыхаемым.
Таблица 3
Содержание газов (в %) во вдыхаемом, выдыхаемом и альвеолярном
воздухе
| Воздух | Кислород | Углекислый газ | Азот |
| Вдыхаемый Выдыхаемый Альвеолярный | 20, 94 16, 3 14, 4 | 0, 03 4, 0 5, 6 | 79, 03 79, 7 80, 0 |
1 Содержание газов во вдыхаемом, выдыхаемом и альвеолярном воздухе обычно приводят из расчета сухого газа, не содержащего паров воды. Название «азот» объединяет содержание азота и всех инертных газов, поскольку все они не участвуют в газообмене.
Выдыхаемый воздух содержит 16, 3% кислорода, но в нем увеличено содержание углекислого газа до 4, 0%- Такое изменение состава воздуха обусловлено тем, что кислород диффундирует из альвеол в капилляры легких, а из крови диффундирует в легкие углекислый газ.
Альвеолярный воздух отличается от выдыхаемого несколько меньшим содержанием кислорода (14, 4%) и несколько большим содержанием углекислого газа (5, 6%). Это объясняется тем, что после газообмена в альвеолах воздух проходит через воздухоносные пути, смешивается с воздухом вредного пространства, в котором не происходит диффузии газов в кровь и из крови.
Диффузия газов из альвеолярного воздуха в гкровь и, наоборот, из крови в альвеолярный воздух определяется соотношением парциальных давлений газов в воздухе и в крови. Парциальное давление газов в крови называется их напряжением (Ро2, Рсо2) и оно также пропорционально содержанию данного газа в крови. Как указывалось выше, диффундировать в жидкость будет тот газ, напряжение которого в жидкости меньше, чем в смеси газов над жидкостью. И наоборот, из жидкости диффундирует тот газ, напряжение которого в ней выше, чем в смеси газов над нею. Данный процесс продолжается до тех пор, пока не устанавливается равновесие между парциальным давлением газа в смеси и напряжением его в жидкости.
Кроме разности в парциальном давлении, растворение газа в жидкости определяется так называемым коэффициентом растворимости газа в жидкости. Коэффициентом растворимости называют то количество газа, которое может быть растворено в 1 мл воды, при давлении 760 мм рт. ст. Коэффициент растворимости зависит от свойства газа, объема и давления газа над жидкостью, температуры жидкости и от количества распространенных в ней веществ.
Альвеолярный воздух непосредственно не соприкасается с кровью, так как отделен от нее тканевыми мембранами. Однако анатомо-физиологические особенности легких создают исключительно благоприятные условия для газообмена. Респираторный аппарат представлен 300 млн. альвеол, соприкасающихся приблизительно с таким же количеством капилляров. Общая поверхность альвеол составляет при вдохе 100 — 120 м2, а толщина легочной мембраны, по данным электронно-микроскопических исследований, равна всего 0, 3—2, 0 мкм. При таких условиях разница между парциальным давлением кислорода и углекислого газа в альвеолярном воздухе и напряжением этих газов в артериальной и венозной крови капилляров легких (табл. 4) вполне достаточна для быстрой диффузии газов.
Из табл. 4 видно, что разность между парциальным давлением кислорода в альвеолярном воздухе и напряжением его в венозной крови капилляров легких равна 110—40 = 70 мм рт. ст., что приводит к диффузии кислорода из альвеолярного воздуха в венозную кровь. Разность между напряжением углекислого газа в венозной крови и парциальным давлением в альвеолярном воздухе равна 47—40 = 7 мм рт. ст., что вызывает диффузию углекислого газа из крови в легкие.
Установлено, что при разности напряжения кислорода в 1 мм рт. ст. в кровь может поступить 15—30 мл кислорода в минуту. Так как средняя величина потребности в кислороде у человека в покое составляет около 250—300 мл в минуту, то при разности давления этого газа в 67—70 мм рт. ст. количество поступающего кислорода более чем достаточно для насыщения крови кислородом.
Оказалось, что скорость диффузии углекислого газа из венозной крови в альвеолярный воздух значительно больше, чем кислорода. Поэтому разность напряжения СО2 7 мм рт. ст. между той и другой средой вполне достаточна для удаления этого газа из венозной крови.
' Таким образом, разница в напряжении кислорода и углекислого газа в альвеолярном воздухе,, в артериальной и венозной крови полностью обеспечивает диффузию необходимого для организма количества кислорода и удаление из крови углекислого газа.
Транспорт газов кровью. Поступившие из альвеолярного воздуха в кровь кислород, а из тканей углекислый газ переносятся кровью, первый из них в ткани, а второй в легкие пере
. Кислород и углекислый газ мало растворимы в плазме крови. Из 100 мл артериальной крови можно выделить 19 мл кислорода, но только 0, 3 мл его находится в растворенном состоянии. Из 100 мл венозной крови можно извлечь 55—58 дол углекислого газа, но только 2, 5 мл его растворено в плазме. Следовательно, после диффузии кислорода и углекислого газа в кровь они не только растворяются в плазме, но и химически связываются с составными частями крови и преимущественно в таком виде ею транспортируются.
Перенос кислорода кровью. Кислород из альвеолярного воздуха диффундирует в плазму, а из нее в эритроциты, где взаимодействует с гемоглобином. Гемоглобин (НЬ) образует с кислородом непрочное, легко диссоциирующее соединение— оксигемоглобин (НЬСЬ). Связывание гемоглобином кислорода зависит от напряжения последнего в крови, но эта зависимость не является прямо пропорциональной. Гемоглобин обладает свойством очень быстро и энергично связывать кислород даже при его низком парциальном давлении, что имеет весьма важное физиологическое значение. Так, например, в растворах гемоглобина на дистиллированной воде при парциальном давлении кислорода 10 мм рт. ст. 55% гемоглобина превращается в оксигемоглобин, а при давлении в 40 мм рт. ст. уже 84% гемоглобина насыщено кислородом.
В нормальных условиях, т. е. при парциальном давлении кислорода в альвеолах в 107—110 мм рт. ст. и напряжении СО2, равном 40 мм рт. ст., 98—99% гемоглобина превращается в оксигемоглобин. Такое интенсивное насыщение венозной крови кислородом обеспечивает его быстрое поступление в кровяное русло и перенос к тканям в достаточном количестве для удовлетворения их потребностей. В тканях оксигемоглобин легко отдает кислород и превращается в восстановленный гемоглобин (ННЬ), который вновь транспортируется к. легким. В тканевых капиллярах кислород легко отщепляется от оксигемоглобина, так как здесь имеется высокое напряжение углекислого газа, который снижает способность гемоглобина связывать кислород.
Таким образом, кислород, поступая из альвеолярного воздуха в кровь, в силу разности парциального давления, частично растворяется в плазме, но главным образом входит в соединение с гемоглобином и в виде оксигемоглобина переносится к тканям.
Перенос углекислого газа кровью. Углекислый газ поступает в кровь из тканей. В тканевых капиллярах растворяющийся в плазме углекислый газ диффундирует в эритроциты, где под влиянием фермента карбоангидразы тут же превращается в углекислоту в соответствии с реакцией СО2+Н2О: 5=ьН2СОз. Благодаря связыванию СО2 все новые количества его поступают в эритроциты. Угольная кислота диссоциирует


Рис. 64. Процессы, происходящие Эритроцит
в эритроците при поглощении или отдаче кровью кислорода и угле-■ кислого газа (схема).
в эритроцитах на ионы Н+ и НСОз". Так как мембрана эритроцитов проницаема для анионов, то ионы НСО3~ диффундируют в плазму. Вместо иона НСО3~ в эритроциты поступают ионы С1~. В плазме благодаря переходу ионов хлора в эритроциты освобождаются ионы натрия, с которыми соединяются ионы НСОз", образуя NaHCO3. Освобождающийся Н+ — ион угольной кислоты в эритроцитах связывается с гемоглобином, и образуется восстановленный гемоглобин. Одновременно с поступлением углекислого газа в эритроциты происходит отдача кислорода окси-
гемоглобином, так как угольная кислота вытесняет из гемоглобина ионы калия, соединяется с ними, образует бикарбонат калия и освобождает кислород. Процесс можно представить как следующую реакцию:
КНЬО2 + Н2СО3 = ННЬ + О2 + КНСО3.
Освобожденный кислород поступает в ткани.
Кроме того, СО2 (приблизительно 8—10% от общего количества,) соединяется с гемоглобином, образуя карбгемо-глобин. Выяснено, что СО2 присоединяется к аминной группе гемоглобина путем так называемой карбаминовой связи.
Таким образом, углекислый газ, поступающий в кровь из тканей, переносится кровью главным образом в виде бикарбонатов и частично в виде карбгемоглобина (рис. 64).
Диффузия углекислого газа в легочных капиллярах. Углекислый газ диффундирует, как указывалось выше, из крови легочных капилляров в альвеолярный воздух вследствие более низкого его парциального давления в альвеолярном воздухе по сравнению с напряжением в венозной крови. Одновременно из альвеолярного воздуха в кровь поступает кислород и в эритроцитах образуется оксигемоглобин. Оксигемоглобин представляет собой кислоту, более сильно диссоциирующую, чем угольная кислота. Поэтому он вытесняет углекис-
лоту из бикарбоната калия по реакции:
ННЬ + О2
= КНЬО2 + Н2СО3.
Угольная кислота расщепляется той же карбоангидразой на СО2 и Н2О, так как этот фермент в зависимости от состава среды катализирует процесс в ту или другую сторону. Углекислый газ диффундирует в альвеолярный воздух, концентрация НСО3~ в крови падает. Это приводит к тому, что карбгемоглобин так же распадается на гемоглобин и СОг. Последний диффундирует в альвеолярный воздух.
Обмен газов между кровью и тканями. Артериальная кровь в тканевых капиллярах отдает кислород и поглощает углекислоту. Диффузия кислорода из крови в ткани обусловлена разностью парциального давления этого газа в крови, где оно высокое, и в протоплазме клеток, где оно низкое. Клетки весьма энергично потребляют кислород, поэтому его напряжение в них может быть равно нулю. В тканевой жидкости оно составляет 20—40 мм рт. ст., а в артериальной крови 102 мм рт. ст., в результате чего кислород непрерывно поступает в тканевую жидкость, а из нее в клетки. Одновременно из клеток через тканевую жидкость в кровь поступает углекислый газ. Его напряжение в клетках может достигать 60 мм рт. ст., в тканевой жидкости оно колеблется и в среднем составляет 46 мм рт. ст., а в артериальной крови — 40 мм рт. ст.
РЕГУЛЯЦИЯ ДЫХАНИЯ
Регуляция дыхания осуществляется рефлекторными и гуморальными механизмами. Оба этих механизма обеспечивают ритмический характер дыхания и изменение его интенсивности, приспосабливая организм к различным условиям окружающей и внутренней среды.
Дыхательный центр и его автоматия. Дыхательным центром называют совокупность специализированных нервных клеток, расположенных в различных отделах центральной нервной системы, которые обеспечивают координированное ритмическое дыхание. Еще в начале XIX века было обнаружено, что возбуждение, вызывающее сокращение дыхательных мышц, возникает в продолговатом мозге. При перерезке головного мозга ниже продолговатого дыхание останавливается и животные погибают. Группы нервных клеток, расположенные в продолговатом мозге, обеспечивающие ритмические.сокращения дыхательных мышц, составляют совершенно необходимый для жизнедеятельности центр дыхания.
Н. А. Миславский в 1885 г. путем раздражения и разрушения отдельных участков продолговатого мозга установил, что дыхательный центр расположен в ретикулярной формации

Рис. 65. Нервные связи дыхательного центра (схема).
/ — инспираторный центр; 2 — центр пневмотаксиса; 3 — экспираторный центр; 4 — механорецепторы легкого. После перерезки по линиям / и // в отдельности ритмическая деятельность дыхательного центра сохраняется. При одновременной перерезке происходит остановка дыхания в фазе выдоха.
продолговатого мозга, в области дна IV желудочка. Центр является парным: от групп нервных клеток в правой половине продолговатого мозга импульсы поступают к дыхательным мышцам правой половины тела, от групп клеток в левой половине — к мышцам левой половины тела.
Дыхательный центр состоит из центра вдоха, или инспираторного центра, и центра выдоха, или экспираторного центра. В верхней части ~ моста мозга (варолиев мост) —одного из отделов заднего мозга, расположенного непосредственно над продолговатым мозгом, находится так называемый центр пневмотаксиса, который также принимает участие в регуляции дыхания.
Импульсы, возникающие в инспираторном центре, распространяются из головного мозга по нисходящим двигательным путям и доходят до двигательных нейронов, расположенных в передних рогах шейных и грудных сегментов спинного мозга. От двигательных нейронов III—IV шейных сегментов отходят аксоны, которые образуют диафрагмальные нервы, снабжающие мышцы диафрагмы. От нервных клеток, расположенных в передних рогах грудного отдела спинного мозга, отходят межреберные нервы, иннервирующие межреберные мышцы.
Для нормальной деятельности дыхательного центра необходима постоянная информация о состоянии внутренней среды организма и самих органов дыхания. Мотонейроны спинного мозга получают от проприорецепторов мышц грудной клетки сигналы о степени растяжения этих мышц при вдохе. Эти сигналы могут изменять число вовлеченных в активность мотонейронов, а следовательно, определяют особенности дыхания. Тем самым осуществляется регуляция дыхания уже на уровне спинного мозга (рис. 65).
Афферентные импульсы бульбарный дыхательный центр получает от механорецепторов легких, дыхательных путей и дыхательных мышц, от хемо- и прессорецепторов сосудистых рефлексогенных зон. Особенно важное значение для регуляции дыхания имеет информация, поступающая от механорецепторов легких по чувствительным нервным волокнам, проходящим в блуждающих нервах (см. ниже).
Дыхательному центру свойственна автоматия, т. е. способность генерировать ритмически импульсы без поступления к нему каких-либо других возбуждений. Ритмическая активность нейронов дыхательного центра сохраняется даже после перерезки всех идущих к нему афферентных нервов. Автоматическое ритмическое возбуждение дыхательного центра обусловлено протекающими в нем процессами обмена веществ и его высокой чувствительностью к напряжению углекислоты. Углекислота всегда содержится в крови и является наиболее сильным раздражителем нейронов дыхательного центра.
Рефлекторная регуляция дыхания. Вдох и выдох обеспечиваются следующими нервными процессами. Благодаря ав-томатии и раздражающему действию углекислоты в инспира-торном центре возникают нервные импульсы, которые, распространяясь по эфферентным путям, приводят к сокращению дыхательных мышц, в результате чего происходит вдох. Растяжение легких вызывает возбуждение механорецепторов, находящихся в стенках альвеол. Импульсы этих рецепторов распространяются по чувствительным нервным волокнам блуждающего нерва к нейронам экспираторного центра и возбуждают его. Одновременно импульсы от инспираторных нейронов поступают в центры пневмотаксиса, а от него приходят к нейронам экспираторного центра и тоже возбуждают его. Возникшее возбуждение экспираторного центра реципрокно тормозит инспираторный центр и импульсация к дыхательным мышцам прекращается. В результате легкие спадаются, происходит выдох. Благодаря этому возбуждение механорецепторов легких и экспираторного центра прекращается. Тормозящее действие нейронов экспираторного центра на инспираторный центр также прекращается. Тогда вновь наступает возбуждение инспираторных нейронов и происходит новый вдох.
Имеется ряд доказательств существования именно такого механизма рефлекторной регуляции дыхания. Например, после перерезки блуждающих нервов дыхание становится резко замедленным и глубоким. В периферическом отрезке блуждающего нерва во время вдоха регистрируются потенциалы действия, доказывающие наличие потока импульсов со стороны легких.
Таким образом, каждый вдох влечет за собой выдох, а выдох стимулирует вдох. В этом и заключается рефлекторная саморегуляция дыхания. Она совершается, как видно из изложенного выше, по принципу автоматического регулирования с механизмом обратной связи. Иначе говоря, она обусловлена взаимодействием между регулирующей (дыхательный центр) и регулируемой (дыхательные мышцы и легкие) системами.
В рефлекторной регуляции принимают участие и другие рецепторные зоны. Так, при выдохе возбуждаются механоре-

Рис. 66. Схема опыта Фредерика с перекрестным кровообращением.
цепторы дыхательных мышц, что рефлекторно стимулирует вдох. При повышении системного артериального давления раздражение барорецепторов каротидного синуса вызывает задержку дыхания. Понижение системного артериального давления ведет к усилению дыхания.
Гуморальная регуляция дыхания. Постоянно изменяющиеся условия окружающей среды, различное физиологическое состояние человека (покой, работа, эмоции и др.) требуют приспособления дыхания по ритму и интенсивности для обеспечения потребности организма в кислороде. Кроме рефлекторной регуляции, дыхание существенно изменяется под влиянием гуморальных факторов. Специфическим гуморальным регулятором дыхательных движений является напряжение СОг, который, накапливаясь в крови, вызывает возбуждение дыхательного центра (рис. 66).
В опыте Фредерика у двух собак, находящихся под наркозом, перерезают и соединяют их сонные артерии и отдельно яремные вены. После такого соединения сосудов и зажатия других сосудов шеи голова первой собаки снабжается кровью от туловища второй собаки, а голова второй — от туловища первой.
Если у одной из этих собак зажать трахею (вызвать удушение), то через некоторое время у нее происходит остановка дыхания, а у второй собаки возникает резкое усиление дыхания (одышка). Последнее объясняется тем, что зажатие трахеи у первой собаки приводит к накоплению СО2 и уменьшению О2 в крови ее туловища. Так как кровь первой собаки поступает к голове второй, то С02 раздражает ее дыхательный Центр и наступает усиленное дыхание.
Остановка дыхания, возникающая у первой собаки, называется апноэ. Усиление и учащение дыхания у второй собаки при накоплении СО2 в крови называется гиперпноэ. Установлено, что увеличение содержания СО2 в альвеолах на 0, 2% ведет к увеличению вентиляции легких на 100%- Уменьшение СО2 в альвеолярном воздухе, а следовательно, понижение его напряжения в крови приводят к ослаблению активности дыхательного центра. Действительно, после усиленного дыхания на некоторое время наступает остановка дыхания. Предварительной усиленной вентиляцией легких пользуются ныряльщики, если им нужно провести под водой 2—3 мин. При обычном спокойном дыхании произвольно задержать дыхание можно только на 40—60 с.
Механизм возбуждающего действия СО2 на дыхательный дентр. Экспериментально было доказано, что всякое увеличение концентрации водородных ионов в крови приводит к усилению дыхания. Поэтому считали, что и при увеличении концентрации в крови водородных ионов, образующихся при диссоциации угольной кислоты, последние проникают в нервные клетки дыхательного центра и возбуждают их. Однако в дальнейшем было показано, что угольная кислота является специфическим раздражителем центра дыхания, так как оказывает на него более сильное возбуждающее действие, чем другие кислоты. Происходит это потому, что углекислый газ легче, чем Н+-ионы, проникает в нейроны дыхательного центра. После диффузии •СО2 в клетках образуется Н2СО3, от которой отщепляются Н+-ионы, возбуждающие нейроны. Сейчас уточнено, -что повышение напряжения СО2 и концентрации Н+-ионов возбуждает центральные нейроны, которые обладают свойствами хеморецепторов. Последние находятся вблизи дыхатель-: ного центра на вентролатеральной поверхности продолговато-то мозга в месте выхода IX и X пар черепных нервов. Раздражителем для них является СО2 спинномозговой жидкости. При воздействии углекислого газа на эти нейроны происходит.стимуляция дыхания; разрушение данных нейронов ведет к прекращению дыхания.
Кроме чувствительных к СО2 и Н+-ионам рецепторов, в -продолговатом мозге имеются аналогичные хеморецепторы, расположенные в рефлексогенных зонах дуги аорты и каро-тидного синуса. Увеличение концентрации Н+-ионов и напряжения СО2 в крови также возбуждает эти рецепторы, что -рефлекторно усиливает дыхательные движения. В этом же направлении оказывает влияние на дыхание пониженное напряжение кислорода в крови.
Таким образом, увеличение напряжения СО2 и концентрации Н+-ИОНОВ в крови регулирует дыхание путем непосредственного влияния этих химических факторов на дыхательный центр и рефлекторно вследствие возбуждения хеморецепторов рефлексогенной зоны каротидного синуса. То и другое усиливают дыхательные движения и ускоряют их ритм. Благодаря усиленному дыханию ускоряется выделение углекислого газа из крови в легкие, увеличивается поступление кислорода в кровь. В этом заключается сущность гуморальной саморегуляции дыхания в соответствии с принципом обратной связи. Здесь деятельность регулирующего аппарата определяется состоянием регулируемого им процесса: накопление в крови углекислоты и недостаток в ней кислорода вызывают возбуждение дыхательного центра, что приводит к усиленному удалению СО2 и поглощению кислорода.
Роль коры полушарий головного мозга в регуляции дыхания. После разрушения дыхательного центра в продолговатом мозге дыхание прекращается, так как этот центр является необходимым для дыхательных движений. Однако в регуляции дыхания принимают участие центры, расположенные в коре полушарий головного мозга. Влияние коры головного мозга на дыхание видно из того, что можно произвольно изменить ритм и глубину дыхания и даже задержать дыхание на некоторое время. При различных эмоциональных реакциях изменяется ритм и глубина дыхания. Можно выработать условный рефлекс на дыхание. Условнорефлекторными механизмами объясняются изменение дыхания перед стартом у спортсмена, особенности дыхания во время речи, пения и в других ситуациях. В течение жизни образуется множество условных дыхательных рефлексов. Этим путем обеспечивается более точное приспособление дыхания к потребностям организма.
Защитные дыхательные рефлексы. Эти рефлексы возникают при раздражении слизистой оболочки дыхательных путей вредными веществами. Защитные дыхательные рефлексы либо препятствуют попаданию вредных веществ в органы дыхания, либо способствуют удалению их из организма. Так, например, при вдыхании аммиака у человека наступает остановка дыхания. Одновременно происходит рефлекторное сужение просвета бронхов и полное закрытие голосовой щели. Благодаря данной реакции вредные вещества не попадают в легкие и организм предохраняется от воздействия летучих ядовитых веществ.
Защитные рефлексы лежат в основе (кашля и чиханья, которые возникают при раздражении слизью, пылью и попадающими в дыхательные пути химическими раздражителями и инородными телами. Перед кашлем человек производит вдох и затем голосовая щель рефлекторно закрывается. Благодаря последующему резкому сокращению дыхательных мышц и раскрытию голосовой щели воздух выталкивается из легких. Сильная струя воздуха увлекает пыль, слизь, инородные тела наружу. Перед чиханьем также происходит сильный вдох, голосовая щель закрывается, мягкое небо поднимается и закрывается носовой ход. Затем, при открывшейся голосовой щели, происходит сильный выдох через нос с характерным звуком и удалением веществ, раздражающих слизистую оболочку.
ДЫХАНИЕ ПРИ МЫШЕЧНОЙ РАБОТЕ
У человека в покое минутный объем дыхания составляет б—8 л воздуха. При мышечной деятельности дыхание учащается, глубина дыхательных движений увеличивается. При напряженной работе объем легочной вентиляции возрастает до 50—60 л в минуту, а у тренированных людей еще больше (до 100—120 л). Такое усиление дыхания обусловлено повышенной потребностью организма в кислороде во время работы и является компонентом общей приспособительной реакции организма к интенсивной мышечной деятельности. Раньше считали, что причиной повышения минутного объема дыхания при мышечной работе является главным образом увеличение напряжения ССЬ в крови и возбуждение в связи с этим центра дыхания.
В настоящее время полагают, что важнейшим фактором, стимулирующим дыхательный центр, являются и афферентные сигналы, идущие от проприорецепторов работающих мышц. Тем самым подчеркивается важное значение рефлекторных каналов регуляции нейронов дыхательного центра. В покое потребление кислорода человеком составляет 250—350 мл в минуту, при мышечной работе оно резко увеличивается и может достигать 4500—5000 мл.
Повышенная потребность организма в кислороде приводит к изменению состояния других органов и систем. При физической работе усиливается работа сердца. За счет увеличения систолического объема и частоты сердцебиений возрастает в.2—3 раза минутный объем крови (до 10—12 л), а у тренированных лиц он может достигать 20—25 л. Через сосуды сердца при такой нагрузке протекает в 3 раза больше крови, чем при его обычной работе. В крови возрастает количество гемоглобина за счет увеличения числа эритроцитов благодаря выходу депонированной крови из селезенки и других органов. Увеличение количества гемоглобина ведет к повышению кислородной емкости крови. Во время работы происходит расширение сосудов в мышцах, открываются дополнительные капилляры, суженные в период покоя. Наступает так называемая рабочая гиперемия благодаря рефлекторным влияниям и образованию во время работы мышц местных физиологически активных веществ. Происходит перераспределение крови в организме, которая притекает в большем количестве к работающим органам, чем к находящимся в покое. При мышечной работе образуются в избыточном количестве молочная и угольная кислоты, мочевина, аммиак и другие соединения. Эти вещества удаляются почками и потовыми железами.
В процессе мышечной работы кровоснабжение почек рефлек-торно уменьшается и мочеобразование задерживается. Однако деятельность потовых желез усиливается в результате рефлекторных влияний и повышения температуры тела. Во время мышечной работы возрастает как образование, так и отдача тепла. Температура тела повышается на 1—1, 5°С, что ускоряет окислительно-восстановительные процессы в тканях и улучшает работоспособность организма.
Таким образом, во время мышечной работы все вышеназванные вегетативные реакции направлены на лучшее обеспечение организма кислородом и энергией в соответствии с возросшими потребностями.
ФУНКЦИОНАЛЬНАЯ СИСТЕМА ВНЕШНЕГО ДЫХАНИЯ
В процессе эволюции сформировались мощные гомеостати-ческие механизмы, обеспечивающие нормальное снабжение тканей кислородом и удаление из них углекислого газа, поскольку от состояния газообмена зависит существование организма. Физиологическое значение кислорода и углекислого газа огромно. Эти газы, с одной стороны, являются непосредственными участниками тканевого дыхания и выработки энергии в митохондриях клеток, а с другой — важнейшими регулируемыми параметрами гомеостаза в организме. В обеспечении тканей кислородом и удалении из них углекислоты принимают участие система внешнего дыхания и система крови и кровообращения, которые тесно взаимодействуют. При этом функциональная система внешнего дыхания играет ведущую роль в поддержании постоянства газового состава крови. Данная система включает совокупность дыхательных нейронов, расположенных на разных уровнях центральной нервной системы, афферентные и эфферентные нервные образования, обеспечивающие связь дыхательных нейронов с исполнительным аппаратом внешнего дыхания (дыхательные пути, легкие, плевра, диафрагма, межреберные мышцы и др.).
Изменение содержания кислорода и углекислого газа в окружающей среде или изменение потребления кислорода и выделения углекислого газа тканями, например при физической нагрузке, отражается на напряжении данных газов в крови, а это в свою очередь немедленно перестраивает деятельность функциональной системы внешнего дыхания. Указанная система, как и все другие гомеостатические системы, функционирует в соответствии с важнейшим принципом саморегуляции — отклонение констант внутренней среды от нормы является стимулом возвращения их к норме. Таким образом, полезным результатом функциональной системы внешнего дыхания является сохранение постоянства газового состава крови— напряжения кислорода и углекислого газа.
В случае снижения Ро2 артериальной крови (гипоксемия) происходит стимуляция хеморецепторов синокаротидной зоны (гипоксические рецепторы), в результате чего усиливаются побуждающие афферентные влияния, идущие к нейронам дыхательного центра. Стимулирующее действие гипоксемии на дыхательный центр происходит рефлекторно, так как непосредственно дефицит кислорода угнетает нейроны дыхательного центра. Напротив, повышение напряжения углекислого газа в крови (гиперкапния), действуя на нейроны дыхательного центра рефлекторно и непосредственно, вызывает их возбуждение. Вероятно, углекислота стимулирует дыхательный центр в результате возбуждения периферических и центральных хеморецепторов. Следствием стимулирующего влияния гипоксемии и гиперкапнии на дыхательный центр является увеличение частоты, а иногда и глубины дыхания, что приводит к повышению минутного объема дыхания и стабилизации газового состава крови. Выравнивание газового состава крови регистрируется теми же хеморецепторами сосудистого русла и мозга, в результате чего ослабляются побуждающие афферентные влияния на дыхательный центр и функциональная система внешнего дыхания нормализует свою деятельность, так как полезный результат достигнут.
Регуляция функциональной системы внешнего дыхания осуществляется не тблько по газовым, но и по негазовым параметрам. К нейронам дыхательного центра поступают многочисленные нервные сигналы, идущие от корковых нейронов и изменяющие легочную вентиляцию (речь, эмоции и произвольные движения), от проприоцепторов мышц, болевых рецепторов, терморецепторов и др. Указанные нервные влияния могут включать систему внешнего дыхания в более сложные по организации функциональные системы, например поведения, в качестве одного из компонентов. Все приходящие к нейронам дыхательного центра влияния принимают участие в афферентной системе. В процессе афферентного синтеза при использовании механизмов памяти происходит подбор такой афферентации, которая способствует удовлетворению доминирующей потребности организма. Результатом афферентного синтеза является принятие решения, и программа данной системы в форме эфферентных влияний изменяет деятельность исполнительных органов, в том числе аппарата внешнего дыхания. Обратная афферентация, идущая от сосудистых и центральных хеморецепторов, механорецепторов легких, дыхательных мышц и других рецепторных аппаратов, поступает в акцептор результата данной функциональной системы, корригируя ее деятельность и состояние внешнего дыхания в соответствии с текущей потребностью организма в газообмене.
НЕКОТОРЫЕ ЗАБОЛЕВАНИЯ ОРГАНОВ ДЫХАНИЯ
Важнейшая область современной медицины — пульмонология успешно развивается в нашей стране в связи с совершенствованием методов диагностики, профилактики и терапии болезней органов дыхания. К наиболее распространенным заболеваниям, развивающимся в дыхательных путях, легких и плевре, относятся ринит (воспаление слизистой оболочки носа), ларингит, трахеит, бронхит, пневмония (воспаление лег--ких) и плеврит. Воспалительный процесс может распространяться на несколько органов системы внешнего дыхания (трахеобронхопневмония).
Причиной воспалительных процессов в органах дыхания могут быть патогенные микроорганизмы (пневмококки, стафилококки, стрептококки), вирусы (гриппа, аденовирусы), ядовитые газообразные вещества и другие факторы. Предрасполагающими факторами возникновения указанных выше заболеваний являются охлаждение, хронические интоксикации (алкоголь, курение).
Течение воспалительных заболеваний органов дыхания может быть острое и хроническое. При остром течении заболевание развивается быстро и бурно. Оно выражается в усиленной секреции желез слизистых оболочек, вследствие чего наступает насморк и отделяется большое количество мокроты. У больного часто возникают сильный кашель, затрудненное дыхание, боли в грудной клетке. Эти явления могут сопровождаться лихорадкой, общей слабостью, головной болью. Однако при правильном лечении воспалительные процессы органов дыхания длятся недолго. При хроническом течении эти процессы менее выражены, развиваются медленно и держатся долго и упорно.
Открытие и применение антибиотиков и сульфаниламидных препаратов открыли новую эру в терапии заболеваний органов дыхания микробного происхождения.
Бронхиальная астма. Это тяжелое заболевание, в основе которого лежат воспаление бронхолегочного аппарата аллергического происхождения. Причиной бронхиальной астмы являются экзогенные аллергены, т. е. вещества разной природы (чужеродные белки, грибки, пыльца растений, бытовая и производственная пыль, краски и др.), вызывающие в организме образование аллергических антител. Развивающееся при этом состояние называется сенсибилизацией. В развитии некоторых форм бронхиальной астмы важное значение имеет наследственная предрасположенность.
Патогенез бронхиальной астмы сложен. Выделяют три стадии развития бронхиальной астмы. В первую, иммунологическую, стадию (сенсибилизация) при ингаляционном поступлении аллергенов в организме образуются аллергические антитела— реагины, которые фиксируются на клетках слизистых оболочек бронхолегочного аппарата. При повторном попадании того же аллергена на слизистых оболочках происходит взаимодействие аллергенов и реагинов. Во вторую стадию, патохимическую, под влиянием образованных комплексов аллерген —• антитело в слизистой оболочке высвобождаются физиологически активные вещества — медиаторы аллергии: медленнореагирующее вещество анафилаксии (МРВ-А), гистамин, ацетилхолин, кинины и др. В третью, патофизиологическую, стадию под влиянием медиаторов аллергии происходит сокращение бронхиол, увеличение секреции желез слизистых оболочек и их отек. Вследствие таких остро возникающих явлений у больных развивается приступ удушья: экспираторная одышка, свистящее дыхание, отхождение мокроты, тяжелый кашель. В процессе заболевания факторами, вызывающими приступ, могут стать не только аллергены, но и другие воздействия: психогенные факторы, физическое напряжение, охлаждение и др.
Эмфизема легких. Данное заболевание часто является исходом острых и хронических воспалительных процессов бронхолегочного аппарата, бронхиальной астмы и др. В основе эмфиземы лежит периодически возникающее повышение внутри-бронхиального и внутриальвеолярного давления при кашле. Это приводит к истончению и разрыву альвеолярных перегородок, ригидности Грудной клетки и снижению легочной вентиляции, а вместе с нарушением легочного кровообращения — к дыхательной гипоксии.
Плеврит. Это заболевание, как правило, является распространением основного заболевания — туберкулеза легких на плевру. В таких случаях часто в плевральной полости скапливается серозно-фибринозный, гнойный или геморрагический экссудат. Туберкулез органов дыхания в прошлом был одной из основных причин смерти. В настоящее время благодаря эффективным мерам социальной профилактики и специфической терапии заболеваемость туберкулезом органов дыхания значительно сократилась.
|