Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Причины и механизмы гаструляции
|
|
В основе перемещения клеточных пластов, обеспечивающих процесс гаструляции, лежат различные морфогенетические движения и процессы.
· увеличение числа клеток посредством делений
Особенно актуально при эпиболии, когда обеспечивается низкий темп деления на вегетативном полюсе и высокая скорость дробления на анимальном полюсе. При инвагинации также отмечается локальное усиление митотической активности в области бластопора.
· растяжение поверхностных клеток эктодермы
Перестройка многослойной стенки крыши бластулы приводит к началу эпиболии (рис. 7).
 А
А 


Б  В
В 
Рис. 7. Интеркаляция нескольких глубоких слоев клеток (Б, В по Гилберту, 1993):
А – схема процесса; Б – строение крыши бластулы лягушки на стадии средней бластулы; В – тоже на стадии средней гаструлы
В ходе реаранжировки клетки наружного ряда уплощаются, стенка бластулы становится тоньше, а клеточный пласт смещается в сторону формирующегося бластопора.
· конвергенция клеток краевой зоны бластулы
Наблюдается в ходе гаструляции у амфибий (рис. 8).

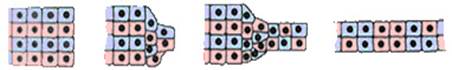
Рис. 8. Конвергентное растяжение (схема)
Представляет собой схождение к центру и удлинение инволюирующего участка бластулы, расположенного непосредственно над областью бластопора. В результате стягивания клеток в более узкую полоску происходит ее растяжение в переднем направлении.
· поляризация клеток
Любое активное изменение формы эпителиального пласта в период гаструляции и нейруляции начинается с того, что его клетки поляризуются, т.е. вытягиваются в перпендикулярном или косом направлении по отношению к поверхности пласта. Поляризация клеток основана на сложных перестройках цитоскелета и клеточной мембраны: сборке микротрубочек и микрофиламентов и их ориентации по длинной оси поляризующейся клетки, а также движениями, так называемых, интегральных белков. В результате этих движений происходит перераспределение ионных каналов и насосов в плазмалемме. Первые концентрируются на внешней стороне клетки, а вторые на боковых и базальных сторонах. Поляризация обуславливает протекание в клетках направленного эндо- и экзоцитоза.
Поляризация затрагивает не одну клетку, а целый клеточный пласт, т.е. поляризация одной клетки побуждает соседнюю к такому же превращению. Этот процесс осуществляется лишь при наличии клеточных контактов и поэтому носит название контактной клеточной поляризации.
Клетки не могут оставаться в состоянии поляризации неограниченно долго. Через определенное время происходит деформация поляризованной клетки, которая уменьшает отношение ее поверхности к объему (сокращение). Этот процесс осуществляется сократительным аппаратом клетки – микрофиламентамии. В ходе последовательных сокращений пласт изгибается, происходит элементарное морфогенетическое перемещение.
· сокращение поляризованных клеток
В результате сокращения апикальных поверхностей поляризован-ных клеток происходит изменение формы всего клеточного пласта образованными ими (рис. 9). Подобные сокращения клеток нейроэкто-дермы играют важную роль в формировании нервной трубки. В данном случае движущей силой является сокращение кольца актиновых микрофиламентов, окружающих апикальные края клеток и связанных с мембраной клетки посредством интегрального белка спектрина.

|
|
|

Рис. 9. Сворачивание пласта клеток (схема):
А, Б – последовательные стадии процесса
· образование колбовидных клеток
В ходе поляризации формируются так называемые колбовидные клетки (рис. 10).
А Б

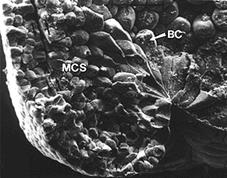
Рис. 10. Колбовидные клетки (по Гилберту, 1993):
А – бокаловидные клетки (тонкая светлая стрелка) до начала инвагинации поверхностного слоя (темная стрелка) внутрь глубоких слоев (толстая светлая стрелка) бластулы; Б – формирующийся бласопор зародыша амфибий (ВС – бокаловидные клетки, МСS – мезодермальные клетки)
Основная часть тела каждой клетки смещена внутрь зародыша, сохраняя контакт с поверхностью посредством узкого цитоплазматического тяжа. Они способны сокращаться, активно перемещаться внутрь, и тянуть за собой остальные клетки пласта. В результате начинает формироваться и постепенно увеличиваться полость архентерона.
Тем не менее оказалось, что после того, как сформировались колбовидные клетки, они не имеют особого отношения к увеличению размеров гастроцеля. Частичное или полное удаление этих клеток не предотвращает инволюцию прилежащих к ним клеток в бластопор.
Главным фактором движения клеток внутрь зародыша является инволюция глубоких клеток краевой зоны бластулы. Эти клетки вворачиваются внутрь и мигрируют к анимальному полюсу по внутренним поверхностям еще неинволюировавших глубоких клеток (рис. 11). Удаление лидирующих глубоких клеток краевой зоны останавливает формирование архентерона.
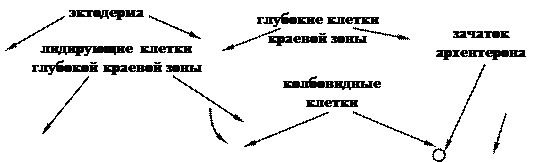 |
|
 Б
Б  В
В 
Рис. 11. Модель клеточных движений в период гаструляции (по Гилберту, 1993):
А, Б, В – последовательные стадии процесса
Таким образом, колбовидные клетки ответственны лишь за образование начального углубления бластопора, а движущая сила инволюции заключена в глубоком слое краевых клеток.
· способность клеток к амебоидным движениям
Адгезивность и подвижность неодинаковы у клеток разных листков. Так клетки эктодермы, контактируя друг с другом, образуют непрерывный пласт, т.е. эпителизируются.
Они способны распространяться над мезодермой и энтодермой. Клетки мезодермы имеют тенденцию инвагинировать в любое находящееся поблизости скопление клеток, а клетки энтодермы относительно неподвижны.
Для гаструляции характерна активность особых групп клеток. Так у морских ежей в начале гаструляции необычайно активны клетки первичной мезенхимы (рис. 12).
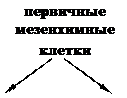 |  | ||
 А
А  Б
Б 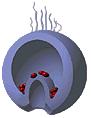 В
В 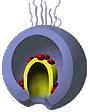 Г
Г 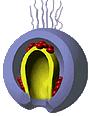
Рис. 12. Поведение мезенхимных клеток при гаструляции у морского ежа (схема):
А – первичные мезенхимные клетки мигрировали внутрь бластоцеля; Б – вторичные мезенхимные клетки попадают внутрь вместе с инвагинирующей первичной кишкой; В, Г – мезенхимные клетки сохраняют свое положение на вершине архентерона, а сокращение их нитевидных отростков, прикрепленных к внутренней поверхности эктодермы, способствует инвагинации архентерона
Они перемещаются от внутренней стенки зародыша в области бластопора в бластоцель, выпускают ламеллоподии (рис. 13) и активно мигрируют к тому месту, где они образуют скелет.



|
|
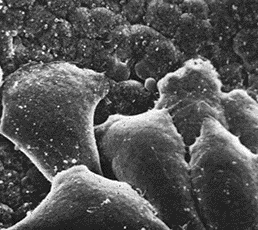
Рис. 13. Мигрирующие при помощи ламеллоподий лидирующие клетки мезодермы зародыша шпорцевой лягушки
Вслед за ними начинает активно инвагинировать остальная часть зародыша.
· роль внеклеточного матрикса
Попав внутрь бластоцеля клетки с помощью ламеллоподий мигрируют по внеклеточному матриксу. Для их успешной миграции важны 2 белка.
Один из них – фибронектин. Это высокомолекулярный гликопротеин (400 кДа), являющийся обычным компонентом базальных мембран.
Исследования показали, что во время гаструляции сродство мигрирующих клеток к данному белку резко возрастает, а сам процесс миграции зависит от концентрации фибронектина (рис. 14).
А  Б
Б 
В  Г
Г 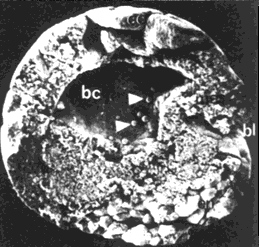
Рис. 14. Роль фибронектинового матрикса в процессах гаструляции у амфибий (по Гилберту, 1993):
А, Б – нормально развившаяся бластула (вид со стороны желточной пробки и поперечный разрез, yp ‑ желточная пробка, ar – архентерон, bc ‑ бластоцель); В, Г – тоже после инъекции синтетического пептида, препятствующего связыванию мигрирующих клеток с фибронектином (bl – бластопор, ес – эктодерма, mes – мезодерма, end – энтодерма)
Клетки связываются с определенным участком фибронектина, содержащим специфическую последовательность из 10 аминокислот.
Вторая важная составляющая часть внеклеточного матрикса – сульфатированные гликопротеины клеточной поверхности мигрирующих клеток.
Важную роль в миграции клеток внутри бластоцеля, особенно у высших позвоночных (птиц и млекопитающих) играют внеклеточные сложные полисахариды. Одним их них является гиалуроновая кислота, линейный полимер глюкуроновой кислоты и N-ацетилглюкозамина. Она накапливается в бластоцеле, покрывая сплошным слоем поверхность вступающих в бластоцель клеток, поддерживая их в диспергированном состоянии за счет способности значительно увеличивать свой объем в воде.
Как ни разнообразны типы гаструляции, существуют общие изменения на клеточном уровне, которые приводят к морфогенетическим перемещениям. Клеточные деления, возникающие в определенном регионе в связи с особыми внешними условиями и воздействиями, создают механические напряжения в пласте. Эти напряжения ведут к возникновению поляризации, которая несет информацию о направлении будущего перемещения, а его реализация, т.е. собственно перемещение, происходит во время сокращения. Морфогенетические движения на протяжении всего развития являются активными. Их источники энергии и исполнительные механизмы находятся внутри того участка, который испытывает ту или иную деформацию.
Факторами организации могут являться механические натяжения тканей зародыша. Первым фактором, обусловившим натяжение эмбриональных тканей, является тургорное давление в полости бластоцеля, растягивающее крышу бластоцеля. Если ослабить его ‑ возникают нарушения в морфогенезе и структуре органов.
Таким образом, в основе большинства формообразовательных процессов лежат клеточные деления, вызывающие, в свою очередь, возникновение механических напряжений в пласте клеток, приводящих к изменению формы зародша.
В регуляции путей развития важную роль играют несколько десятков белков, именуемых факторами роста, которые в организме животных регулируют клеточные деления и дифференцировку, а также возникновение тканей. Подобно нейромедиаторам в нервной системе такие факторы, по-видимому, используются в разных обстоятельствах для передачи различных сигналов между клетками.
Контрольные вопросы
1. Определение гаструляции. Периодизация гаструляции.
2. Типы движений при гаструляции.
3. Телобластический и энтероцельный способы закладки мезодермы.
4. Образование вторичной полости тела (целома).
5. Производные зародышевых листков.
6. Клеточные основы процесса гаструляции.
Литература