Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Состав и функции биологических мембран.
|
|
Структура и функции биологических мембран.
Состав и функции биологических мембран.
Биологическими мембранами называют функциональные структуры клеток, толщина которых составляет несколько молекулярных слоев, ограничивающие цитоплазму и большинство внутриклеточных структур. Они образуют единую внутриклеточную структуру каналов, складок и замкнутых полостей. Толщина биологических мембран редко превышает 10, 0 нм, но вследствие плотной упаковки в них основных молекулярных компонентов (белки и липиды), а также большой общей площади клеточных мембран они составляют обычно более половины массы сухих клеток.
Исследования показали, что основными химическими компонентами мембран являются липиды (≈ 30%), белки (≈ 60%) и углеводы (≈ 10%). Углеводы связаны с молекулами белка (гликопротеины), либо с молекулами липидов (гликолипиды). Минорными компонентами мембран являются нуклеиновые кислоты, полиамины, неорганические ионы.
1.Липиды, входящие в состав мембран, принадлежат к трем классам. Это фосфолипиды, гликолипиды и стероиды.
Фосфолипиды построены по единому плану, их молекула имеет полярную головку и два неполярных хвоста.
В состав головки фосфолипида обязательно входят:
а) остаток спирта (реже аминокислоты);
б) остаток фосфорной кислоты;
в) по вариантам либо:
– глицерин (глицерофосфолипиды),
– многоатомный спирт сфингозин (сфингофосфолипиды).
Неполярные хвосты всех молекул фосфолипидов образуются жирными кислотами, определяющими разнообразие их представительства.
Глицерофосфолипиды мембран растительных клеток содержат в основном пальмитиновую (С16: 1(9)), олеиновую (С18: 1(9)), линолевую (С 18: 2(9, 12)) жирные кислоты. Жирные кислоты с количеством атомов углерода более 20–24 в растительных клетках встречаются редко.
Глицерофосфолипиды мембран животных клеток содержат пальмитиновую, олеиновую, стеариновую (С18: 0) жирные кислоты, а также кислоты (арахидоновая и др.) с числом атомов углерода, превышающим 20.
Обычно из двух жирных кислот, образующих гидрофобный хвост глицерофосфолипида, одна ненасыщенная. Она образует связь со вторым атомом углерода глицерина и одна из ее двойных связей обязательно находится на уровне 9–го атома углерода, считая от головки.
Остаток спирта (аминокислоты) головки глицерофосфолипида связан сложноэфирной связью с фосфорной кислотой. Глицерофосфолипид, в составе которого отсутствует спирт (или аминокислоты), называется фосфатидной кислотой. Фосфатидная кислота является промежуточным продуктом в биосинтезе фосфолипидов и в свободном виде практически не встречается.
В зависимости от того, какой именно спирт (аминокислота) входит в состав головки молекулы, различают несколько классов глицерофосфолипидов.
Самые распространенные глицерофосфолипиды:
– фосфатидилхолин, содержащий спирт холин;
– фосфатидилэтаноламин, содержащий спирт этаноламин;
– фосфатидилинозитол, содержащий циклический спирт инозитол;
– фосфатидилсерин, содержащий остаток гидроксиаминокислоты, – серина.
Примерами сфингофосфолипидов могут служить сфингомиелины, которые в составе полярной головки содержат фосфохолин или фосфоэтаноламин. В составе сфингофосфолипидов одна жирная кислота.
Таким образом, фосфолипиды содержат группировки двух видов – полярные гидрофильные головки и неполярные гидрофобные хвосты. Вследствие этого фосфолипиды обладают амфипатическими (амфифильными) свойствами. При рН = 7 остаток фосфорной кислоты в полярной головке заряжен отрицательно.
2.Белки мембраны и их функции. Доля белка в общей массе мембраны может колебаться в очень широких пределах – от 18 % в миелине до 75 % в митохондриальной мембране.
По месту расположения в мембране белки делят на интегральные и периферические (рис. 4.1).
Интегральные белки являются, как правило, гидрофобными и легко встраиваются в липидный бислой.
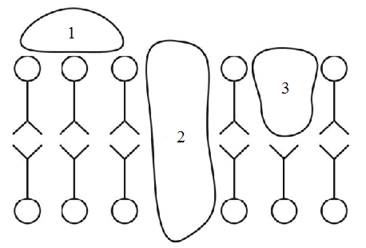
Рис. 4.1. Классификация белков по месту их расположения в мембране:
1 – периферические; 2 – интегральные; 3 – полуинтегральные.
Взаимодействие такого белка с мембраной происходит в несколько стадий. Сначала белок адсорбируется на поверхности бислоя, изменяет свою конформацию, устанавливая гидрофобный контакт с мембраной. Затем происходит внедрение белка в бислой. Глубина внедрения зависит от силы гидрофобного взаимодействия и соотношения гидрофобных и гидрофильных участков на поверхности белковой глобулы. Гидрофильные участки белка взаимодействуют с примембранными слоями по одну или обе стороны мембраны. Фиксация белковой глобулы в мембране происходит благодаря электростатическим и гидрофобным взаимодействиям. Углеводная часть белковых молекул (если она имеется) выступает наружу. Интегральные белки в силу тесной связи с бислоем оказывают на него существенное воздействие: конформационные перестройки белка приводят к изменению состояния липидов, так называемой деформации бислоя.
Периферические белки обладают меньшей глубиной проникновения в липидный бислой и, соответственно, более слабо взаимодействуют с липидами мембраны, оказывая на них гораздо меньшее воздействие, чем интегральные.
Выделяют также промежуточный вид белков – полуинтегральные.
По характеру взаимодействия с мембраной белки делятся на монотопические, битопические, политопические:
– монотопические белки взаимодействуют с поверхностью мембраны (моно – одним из слоев липидов);
– битопические пронизывают мембрану насквозь (би – двумя слоями липидов);
– политопические пронизывают мембрану несколько раз (поли – многократное взаимодействие с липидами).
Первые относятся к периферическим белкам, а вторые и третьи – интегральным.
Белки мембран можно также классифицировать по выполняемой ими функции. В связи с этим выделяют следующие белки:
– структурные белки;
– белки–ферменты;
– белки–рецепторы;
– транспортные белки.
Особую группу составляют белки цитоскелета клетки. Строго говоря, эти белки не являются компонентами мембраны, примыкая к ней с цитоплазматической стороны. Белки цитоскелета входят в состав всех его компонентов: миофиламенты содержат молекулы белка актина; в состав микротрубочек входит белок тубулин, промежуточные филаменты также содержат более полиморфный белковый комплекс. Цитоскелет не только обеспечивает эластичность мембраны, противостоит изменениям объема клетки, но, по-видимому, участвует и в различных внутри- и внеклеточных механизмах регуляции.
3.Важным структурным компонентом мембран является вода. Воду, входящую в состав мембран, подразделяют на группы: связанную, свободную и захваченную воду. Наименьшей подвижностью обладает связанная вода, присутствующая в виде одиночных молекул в углеводородной зоне мембран. Основная часть связанной воды – это вода гидратных оболочек макромолекул. Гидратные оболочки образуются главным образом вокруг полярных частей молекул липидов и белков. Гидратные оболочки основных структурообразующих липидов состоят обычно из 10–12 молекул воды. Эта вода в осмотическом отношении неактивна и неспособна растворять какие – либо вещества.
Иногда в составе связанной воды выделяют слабосвязанную воду. Слабосвязанная вода по подвижности и некоторым другим свойствам занимает промежуточное положение между водой гидратных оболочек и жидкой свободной водой.
Свободная вода входит в состав мембран в виде самостоятельной фазы и обладает подвижностью, как и у жидкой воды.
Захваченная вода обнаружена в центральной части мембран между липидными бислоями. По параметрам подвижности она соответствует жидкой свободной воде, но медленно обменивается с внешней средой из-за физической разобщенности.
Функции биологических мембран. Основные функции (выполняются всеми без исключения мембранами):
1.Компартментализация – участие мембран в образовании изолированных отсеков (компартментов). Мембраны образуют замкнутые структуры, плазматическая мембрана отделяет клетку от окружающей среды.
2.Барьерная функция – препятствие свободной диффузии различных веществ. В результате мембрана регулирует перенос веществ и их содержание в клеточных и субклеточных компартментах.
3.Рецепторная (сигнальная) функция – связывание мембранными рецепторами внеклеточных эффекторов – биологически (физиологически) активных веществ, что приводит к изменению активности регуляторных белков клетки.
4.Ферментативная функция обеспечивается мембранно–связанными белками–ферментами, которые образуют своеобразные конвейеры.
5.Энергетическая – синтез АТФ на внутренних мембранах митохондрий и фотосинтез в мембранах хлоропластов.
Факультативные функции, присущие не всем мембранам:
– транспорт электронов и фосфорилирование в дыхательной цепи (способны выполнять мембраны митохондрий, хлоропластов);
– преобразование внешних стимулов в электрические импульсы;
– проведение биоэлектрических импульсов (эта функция характерна для мембран электровозбудимых клеток: нервной и мышечной ткани);
– высвобождение синаптических нейромедиаторов и пиноцитоз.