Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Тема 11. Молекулярные основы наследственности
|
|
Задания
1. Разобраться в схеме репликации, транскрипции и трансляция генетического материала в клетке.
2. Разобраться в схеме строения гена.
3. Выяснить назначение и механизм процессинга у эукариот.
4. Решить задачи.
Литература
1. Гуляев Г.В. Генетика. – 3-е изд., перераб. и доп. – М.: Колос, 1984. – С. 125-173.
2. Айала Ф., Кайгер Дж. Современная генетика: Пер. с англ. – М.: Мир, 1987. – Т. 1. – С. 88-295.
3. 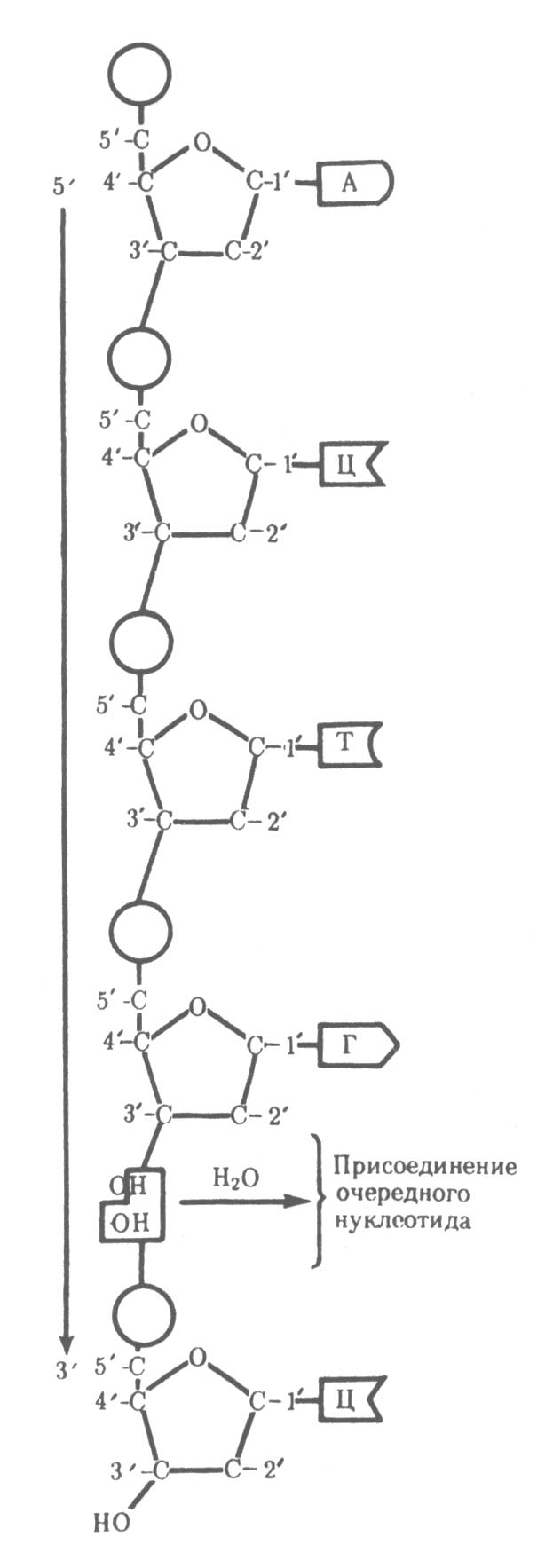
| Рисунок 31 – Схема полинуклеотидной цепи(стрелкой указано направление роста цепи) |
4. Биология / В.Н. Ярыгин, В.И. Васильева, И.Н. Волков, В.В. Синельщикова; Под ред. В.Н. Ярыгина. – 2-е изд., испр. – М.: Высш. шк., 1999. – 448 с.
5. Инге-Вечтомов С.Г. Генетика с основами селекции. – М.: Высш. шк., 1989. – 521 с.
6. Гуляев Г. В. Задачник по генетике. – М.: Колос, 1973. – С. 28-29.

| Рисунок 30 – Схема строения нуклеотида |
Каждая нить молекулы ДНК состоит из нуклеотидов. Нуклеотид, в свою очередь, состоит из пентозного сахара дезоксирибозы, остатка фосфорной кислоты и одного из четырёх азотистых оснований (пуриновые – 
| Рисунок 32 – Схема строения молекулы ДНК(стрелками обозначена антипараллельность нитей) |
Цепи молекулы ДНК соединяются друг с другом водородными связями между их азотистыми основаниями по принципу комплементарности. Аденин одной цепи соединяется двумя водородными связями с тимином другой цепи, а между гуанином и цитозином разных цепей образуются три соединяющих их связи.
Другой важной особенностью объединения двух полинуклеотидных цепей в молекуле ДНК является их антипараллельность: 5 ′ -конец одной цепи находится рядом с 3′ -концом другой, и наоборот.
Таким образом, в структурной организации молекулы ДНК выделяют первичную структуру – полинуклеотидная цепь, вторичную структуру – две комплементарные друг другу и антипараллельные полинуклеотидные цепи, соединённые водородными связями, и третичную структуру – несколько форм трёхмерной спирали (А-, В-, С- и Z-форма).
Одним из основных свойств наследственного материала клетки является его способность к самокопированию – репликации. В процессе репликации на каждой полинуклеотидной цепи материнской молекулы ДНК синтезируется комплементарная ей цепь. В итоге из одной двуцепочечной спирали ДНК образуется две идентичные молекулы ДНК. Такой способ удвоения молекул, при котором каждая дочерняя молекула содержит одну исходную материнскую и одну вновь синтезированную цепь, называют полуконсервативным.
Репликация у эукариот начинается в определённых точках (локус ori) с образования особой структуры – репликационного глаза, где две цепи родительской ДНК ферментом геликазой (хеликазой) отделяются друг от друга, чтобы стать матрицами синтеза комплементарных цепей дочерних молекул. Область расхождения полинуклеотидных цепей в зонах репликации называют репликационными вилками. Фрагмент ДНК от одной точки репликации до другой точки образует единицу репликации – репликон. В процессе синтеза две репликационные вилки, образующие репликационный глаз, перемещают 
| РНК-праймаза |
| Матричная цепь ДНК |
| Затравка |
| Рисунок 33 – Схема синтеза затравки |
Разделённые геликазой нити материнской ДНК связываются с белком SSB, предотвращая преждевременную реассоциацию цепей и их разрушение.

| Рисунок 34 – Снятие напряжения двойной спирали молекулы ДНК с помощью фермента ДНК‑ топоизомераза: А – ДНК-топоизомераза образует ковалентную связь с одной из фосфатных групп ДНК; В– в результате разрыва фосфодиэфирной связи в одной полинуклеотидной цепи вокруг соответствующей ей связи другой цепи осуществляется вращение, которое снимает напряжение, вызванное расхождением двух цепей ДНК в области репликационой вилки; С– после снятия напряжения в спирали ДНК происходит спонтанное отделение ДНК‑ полимеразы, которая перемещается дальше перед репликационной вилкой, и восстанавливается фосфодиэфирная связь в цепи ДНК |
Разделение спирально закрученных цепей родительской ДНК ферментом геликазой вызывает появление супервитков перед репликационной вилкой. Фермент топоизомераза, работая перед репликационной вилкой, разрывает одну из цепей молекулы ДНК, давая ей возможность вращаться вокруг второй цепи и снимая накопившееся напряжение в двойной спирали ДНК (рисунок 34).

| Рисунок 35 – Присоединение ДНК-полимеразой очередного нуклеотида к дочерней цепи ДНК |
Способность ДНК-полимеразы осуществлять сборку полинуклеотидов в направлении 5′ - к 3′ -концу при антипараллельном соединении двух цепей ДНК означает, что процесс репликации должен протекать на них по-разному. Так, если на лидирующей дочерней цепи ДНК, которая синтезируется по матрице, имеющей направление 3′ 5′ , её рост идет непрерывно от 5′ - к 3′ -концу, то другая отстающая дочерняя цепь не может формироваться по ходу репликационной вилки. Поэтому её синтез осуществляется в направлении, обратному ходу репликационной вилки, отрезками протяжённостью (у эукариот) 100-200 нуклеотидов, называемыми фрагментами Оказаки. Синтезу каждого такого фрагмента предшествует образование затравки (см. рисунок 33). Два сформированные фрагмента Оказаки после удаления из них затравки (осуществляется ферментом ДНК-полимераза I) сшиваются ферментом ДНК-лигаза с образованием фосфодиэфирной связи, формируя тем самым отстающую нить (рисунок 36).
Общая схема работы ферментов в репликационной вилке показана на рисунке 37.

| Рисунок 36 – Синтез отстающей дочерней цепи молекулы ДНК |
Известно, что хромосомы эукариотических клеток состоят в основном из ДНК и белков, образуя нуклеопротеиновый комплекс. На долю белков (гистоновые и негистоновые) приходится около 60 % массы всей хромосомы. Гистоны представлены пятью белковыми фракциями: Н1, Н2А, Н2В, Н3, Н4. Являясь положительно заряженными основными белками, они достаточно прочно соединяются с молекулами ДНК, препятствуя считыванию заключённой в ней информации. В этом состоит их регуляторная роль. Кроме того, эти белки обеспечивают пространственную организацию ДНК в хромосомах.
В процессе подготовки клетки к делению интерфазное состояние ядра начинает меняться, приводя к образованию строго специфичного для вида кариотипа хромосом (количество, форма, размер). Основную роль в этом процессе выполняют гистоны.

| Рисунок 38 – Нуклеосомная организация хроматина |
| Рисунок 37 – Схема участия основных белков в процессе репликации ДНК |

| Геликаза |
| Фрагменты Оказаки |
| Материнские цепи молекулы ДНК |
| Топоизомераза |
| Дестабилизирующий белок |
| Лидирующая цепь |
| Отстающая цепь |
Первый уровень организации хроматина – нуклеосомная нить обеспечивается четырьмя видами гистонов Н2А, Н2В, Н3, Н4 (рисунок 38). Они образуют округлую белковую структуру, состоящую из восьми молекул (по две молекулы каждого вида гистонов), на которую спирально накручивается молекула ДНК, состоящая из 146 пар нуклеотидов (п.н.). Свободные от контакта с белковыми телами участки ДНК, включающие от 15 до 100 п.н. (в среднем 60 п.н.), называются связующими или линкерными. Спирально накрученная на гистоновую структуру молекула ДНК вместе с линкерной последовательностью составляет длину около 200 п.н. и называется нуклеосомой.
Молекула ДНК, упакованная в виде нуклеосомной нити, напоминает цепочку бус. Её диаметр с 2 нм приобретает 10-11 нм, а длина уменьшается с 5 до 0, 7 см.
Дальнейшая компактизация нуклеосомной нити обеспечивается гистоном Н1, который, соединяясь с линкерной ДНК и двумя соседними белковыми телами, сближает их друг с другом. В результате образуется элементарная хроматиновая фибрилла, имеющая диаметр 20-30 нм и длину 1, 2 мм.
Следующий уровень структурной организации генетического материала обусловлен укладкой хроматиновой фибриллы в петли (рисунок 39). В результате такой упаковки хроматиновая фибрилла диаметром 20-30 нм преобразуется в структуру диаметром 100-200 нм, называемую интерфазной хромонемой.

| Рисунок 39 – Интерфазная хромонема А – хроматиновая фибрилла с присоединёнными к ней негистоновыми белками; Б – образование петли на участке хроматиновой фибриллы; В – схема петельной организации участка хромосомы |
Отдельные участки интерфазной хромонемы подвергаются дальнейшей компактизации, образуя структурные блоки, объединяемые в дальнейшем в метафазные хромосомы.
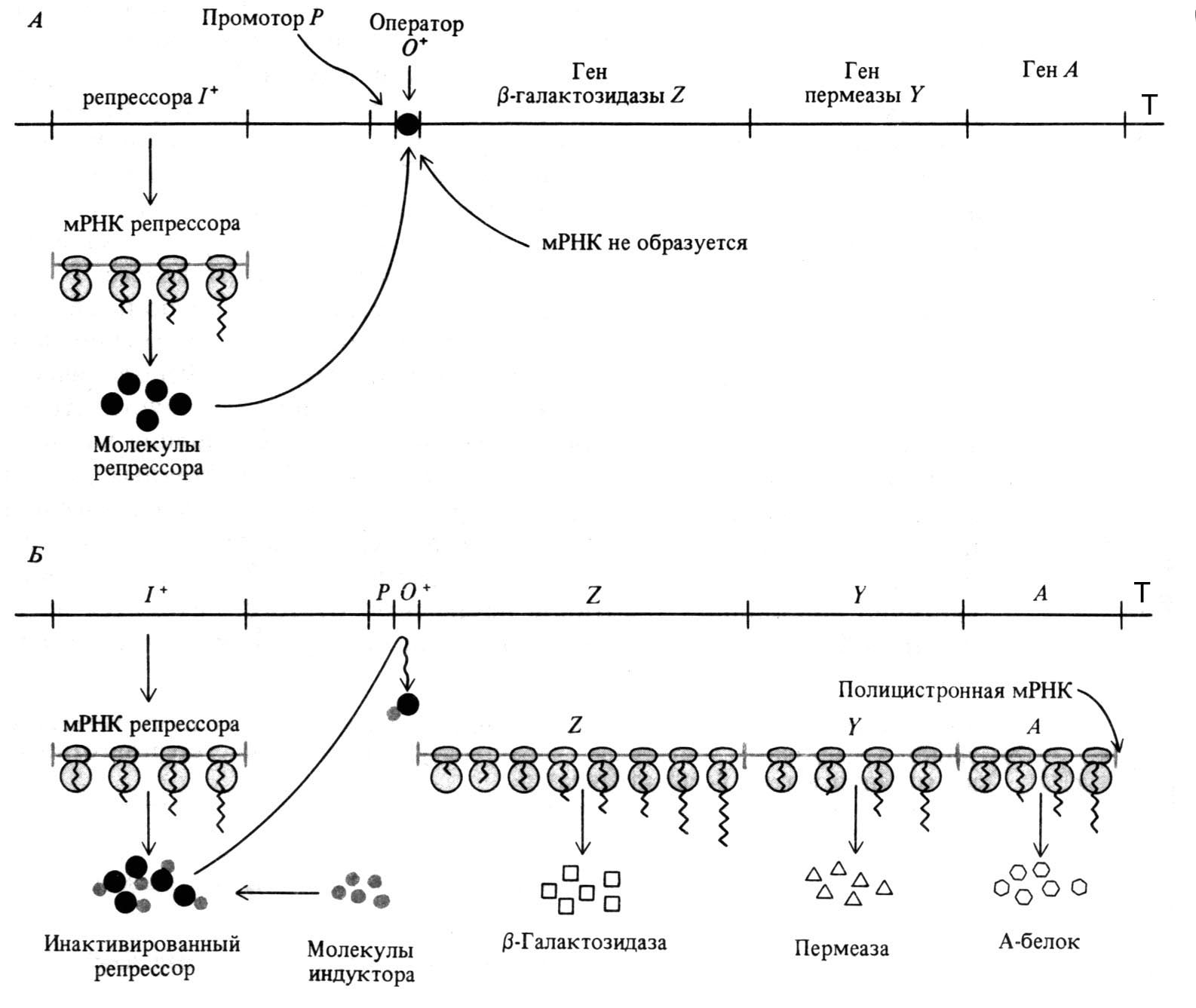
Молекула ДНК имеет значительную протяжённость. Ген – это участок молекулы ДНК протяжённостью около 1000 нуклеотидов. Ген включает в себя не только структурную часть, но и регуляторные последовательности. В 1961 г. Жакоб и Моно выдвинули предположение, что транскрипция генов находится под контролем регуляторного участка, названного оператором (О). Регуляция транскрипции на операторном участке осуществляется репрессором, вырабатываемым геном-индуктором (I). Репрессор, связываясь с оператором, подавляет транскрипцию. Если же репрессор связывается с индуктором, то происходит диссоциация комплекса репрессор-оператор и тем самым делая возможным транскрипцию (рисунок 40).
| Ген |
| Структурная часть |
| Структурная часть |
| Рисунок 40 – Схема взаимодействия регуляторных частей гена: А – репрессор связывается с оператором, препятствуя соединению РНК-полимеразы с промотором и началу транскрипции; Б – индуктор связывается с репрессором, инактивируя его, что даёт возможность РНК‑ полимеразе связаться с промотором и начать транскрипцию. |
Синтез м-РНК (и-РНК) начинается с обнаружения РНК-полимеразой особого участка на молекуле ДНК (промотор), который указывает место начала транскрипции. После присоединения к промотору РНК-полимераза раскручивает виток спирали ДНК и две её цепи расходятся. Сборка рибонуклеотидов в цепь происходит с соблюдением их комплементарности нуклеотидам ДНК, а также антипараллельно по отношению к матричной цепи ДНК. В связи с тем, что РНК-полимераза способна собирать полинуклеотид лишь от 5′ - к 3′ -концу, матрицей для транскрипции может служить только та цепь, которая обращена к ферменту 3′ -концом (эту цепь называют кодогенной).
РНК-полимераза, продвигаясь вдоль кодогенной цепи ДНК, осуществляет понуклеотидное переписывание информации до тех пор, пока не встретит специфическую нуклеотидную последовательность – терминатор. В этом участке РНК-полимераза отделяется как от матрицы ДНК, так и от вновь синтезированной м-РНК. По мере продвижения РНК-полимеразы пройденные ею одноцепочечные участки ДНК вновь объединяются в двойную спираль.
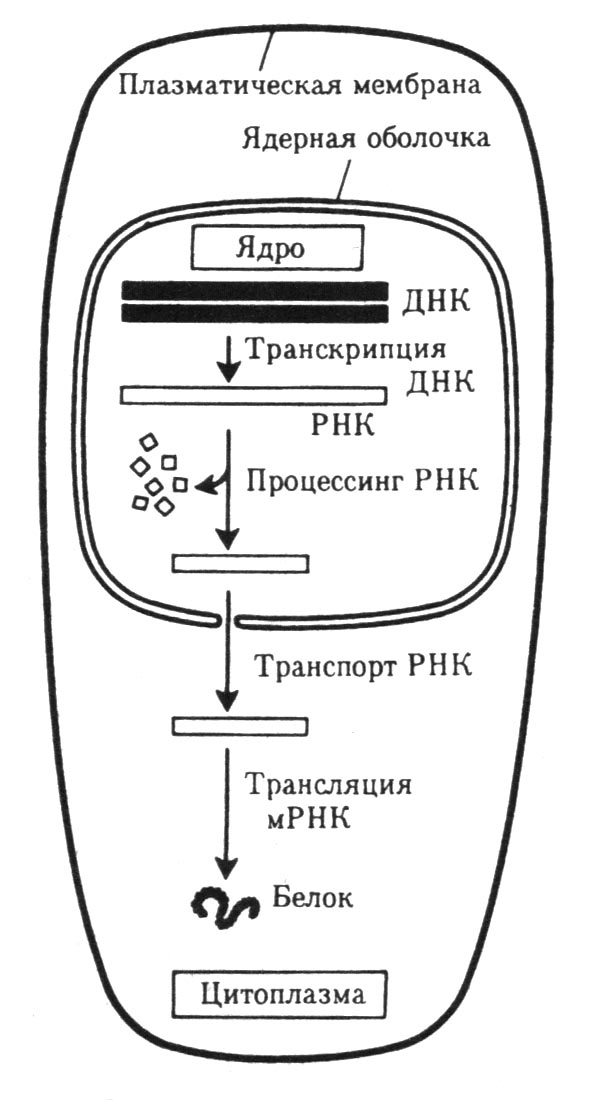
| Рисунок 41 – Схема экспрессии генетической информации в эукариотической клетке |
Процессинг (созревание) гя-РНК предполагает модифицирование первичного транскрипта и удаление из него некодирующих интронных участков с последующим соединением (сплайсинг) кодирующих последовательностей – экзонов. Кроме сплайсинга во время процессинга происходит также модифицирование первичного транскрипта с 5′ -конца путём образования колпачка – кэп, который обеспечивают узнавание молекул м-РНК рибосомами цитоплазмы (рисунок 42).
Кроме того, происходит удаление части нуклеотидов на 3′ -конце первичного транскрипта и присоединение к нему последовательности, состоящей из 100-200 остатков адениловой кислоты (поли-А). Считается, что эта последовательность способствует энергетическому обеспечению дальнейшего процессинга и транспорта зрелой м-РНК из ядра в цитоплазму.
Наряду с кэпированием и полиаденилованием первичного транскрипта происходит удаление неинформативных для данного белка интронных участков, размер которых варьирует от 100 до 10000 нуклеотидов и более. На долю интронов приходится около 80 % всей гя-РНК. Удаление интронов с последующим соединением экзонов осуществляется специальными структурами – малыми ядерными РНК (мя-РНК).

| Первичный транскрипт |
| Модифицирование концевых участков первичного транскрипта |
| Удаление интронных участков |
| Сплайсинг экзонов |
| Поли-А |
| Поли-А |
| Поли-А |
| Зрелая м-РНК |
| Рисунок 42 – Образование зрелой м-РНК эукариот в ходе процессинга: 1 – некодирующие последовательности, 2 – экзоны, 3 – интроны, 4 – кодон-терминатор |
В настоящее время доказана возможность альтернативного сплайсинга, при котором на одном первичном транскрипте могут удаляться разные нуклеотидные последовательности и образовываться различные зрелые м-РНК.
Наследственная информация, записанная с помощью генетического кода, хранится в молекулах ДНК и непосредственного участия в жизнеобеспечении клеток не принимает. Роль посредника в переводе наследственной информации, хранимой в ДНК, в рабочую форму играют рибонуклеиновые кислоты – РНК.
Молекулы РНК состоят из четырёх типов нуклеотидов, содержащих сахар рибозу, фосфат и одно из четырёх азотистых оснований – аденин, гуанин, цитозин, урацил. Всё многообразие РНК, действующих в клетке, можно разделить на три основных вида: м-РНК, т-РНК, р-РНК.
Матричная РНК (м-РНК) синтезируется РНК-полимеразой на ДНК как копия соответствующего гена. Этот процесс называется транскрипцией. Тройки рядом стоящих нуклеотидов м-РНК, шифрующие определённые аминокислоты, называются кодонами (триплеты). Таким образом, последовательность кодонов м-РНК шифрует последовательность аминокислот в пептидной цепи (рисунок 43, таблица 10).
Генетический код характеризуется следующими основными свойствами:
¨ универсальность для живых организмов,
¨ триплетность (состоит из трёх нуклеотидов),
¨ неперекрываемость (азотистые основания одного триплета не входят в состав других соседних триплетов),
¨ вырожденность (одна аминокислота может кодироваться несколькими триплетами).
| Рисунок 43 – Генетический код |

В генноинженерных работах имеется необходимость по аминокислотной последовательности белка определить нуклеотидную последовательность и‑ РНК. Для решения задач на эту тему удобно пользоваться генетическим кодом, представленным в виде таблицы 10.
Таблица 10 – Последовательность нуклеотидов в кодонах и-РНК для разных аминокислот
| Аминокислоты | Кодоны | |||||
| Фенилаланин (фен) | УУУ | УУЦ | ||||
| Лейцин (лей) | УУА | УУГ | ЦУУ | ЦУЦ | ЦУА | ЦУГ |
| Изолейцин (илей) | АУУ | АУЦ | АУА | |||
| Метионин (мет) | АУГ | |||||
| Валин (вал) | ГУУ | ГУЦ | ГУА | ГУГ | ||
| Серин (сер) | УЦУ | УЦЦ | УЦА | УЦГ | АГУ | АГЦ |
| Пролин (про) | ЦЦУ | ЦЦЦ | ЦЦА | ЦЦГ | ||
| Треонин (тре) | АЦУ | АЦЦ | АЦА | АЦГ | ||
| Аланин (ала) | ГЦУ | ГЦЦ | ГЦА | ГЦГ | ||
| Тирозин (тир) | УАУ | УАЦ | ||||
| Гистидин (гис) | ЦАУ | ЦАЦ | ||||
| Глутамин (глн) | ЦАА | ЦАГ | ||||
| Аспарагин (асн) | ААУ | ААЦ | ||||
| Аспарагиновая кислота (асп) | ГАУ | ГАЦ | ||||
| Лизин (лиз) | ААА | ААГ | ||||
| Глутаминовая кислота (глу) | ГАА | ГАГ | ||||
| Цистеин (цис) | УГУ | УГЦ | ||||
| Триптофан (трип) | УГГ | |||||
| Аргинин (арг) | ЦГУ | ЦГЦ | ЦГА | ЦГГ | АГА | АГГ |
| Глицин (гли) | ГГУ | ГГЦ | ГГА | ГГГ | ||
| Охра* | УАА | |||||
| Амбер* | УАГ | |||||
| Опал* | УГА | |||||
| * Примечание: охра, амбер, опал – условные названия терминирующих триплетов (нонсенс-кодоны) |
.
Важная роль в процессе реализации наследственной информации клетки принадлежит транспортной РНК (т-РНК), которая доставляет соответствующие аминокислоты к месту сборки пептидных цепей.
Молекулы т-РНК представляют собой относительно небольшие нуклеотидные последовательности (75-95 нуклеотидов) и комплементарно соединённые в определённых участках. В результате формируется структура, напоминающая по форме лист клевера, в которой выделяют две наиболее важные зоны – акцепторная часть и антикодон.
Акцепторная часть т-РНК состоит из комплементарно соединённых 7 пар оснований и несколько более длинного одиночного участка, заканчивающегося 3′ -концом, к которому присоединяется транспортируемая соответствующая аминокислота (рисунок 44).

| Рисунок 44 – Схема строения т-РНК |
Наряду с функцией точного узнавания определённого кодона в м-РНК молекула т-РНК связывается и доставляет к месту синтеза белка определённую аминокислоту, присоединённую ферментом аминоацил-т-РНК-синтетазы. Этот фермент обладает способностью к пространственному узнаванию, с одной стороны, антикодона т-РНК и, с другой, – соответствующей аминокислоты. Для транспортировки 20 типов аминокислот используются свои транспортные РНК.
Процесс взаимодействия м‑ РНК и т-РНК, обеспечивающий трансляцию информации с языка нуклеотидов на язык аминокислот, осуществляется на рибосомах.
Рибосомы представляют собой сложные комплексы рибосомной РНК (р‑ РНК) и разнообразных белков. Рибосомная РНК является не только структурным компонентом рибосом, но и обеспечивает связывание её с определённой нуклеотидной последовательностью м-РНК, устанавливая начало и рамку считывания при образовании пептидной цепи. Кроме того, они обеспечивают взаимодействие рибосомы с т-РНК (рисунок 45).

| Малая субчастица |
| Большая субчастица |
| (Р-участок) |
| (А-участок) |
| Рисунок 45 – Участки связывания т-РНК и рибосомы |
В ходе трансляции выделяют три фазы: инициацию, элонгацию и терминацию синтеза пептидной цепи.
Фаза инициации заключается в объединении двух находящихся до этого порознь в цитоплазме субъединиц рибосомы на определённом участке м-РНК и присоединение к ней первой аминоацил-т-РНК. Этим также задаётся рамка считывания информации, заключённой в м-РНК.
Фаза элонгации (или удлинения пептида) включает в себя все реакции от момента образования первой пептидной связи до присоединения последней аминокислоты. Она представляет собой циклически повторяющееся специфическое узнавание аминоацил-т-РНК очередного кодона, находящегося в А‑ участке, комплементарное взаимодействие между кодоном и антикодоном.
В А-участке т-РНК располагается таким образом, что её аминокислота находится рядом с аминокислотой Р-участка. С помощью специальных белков между двумя аминокислотами образуется пептидная связь, освобождая т-РНК, находящуюся в Р-участке. В этот момент рибосома передвигается по м-РНК на один шаг (триплет). Так продолжается до тех пор, пока в А-участок рибосомы не поступит кодон-терминатор. Скорость синтеза аминокислот в эукариотических клетках составляет 2 аминокислоты в 1 с.
Фаза терминации (завершения синтеза пептида) связана с узнаванием терминирующих кодонов. В результате завершённая пептидная цепь теряет связь с рибосомой, которая распадается на две субъединицы.