Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Оогенез у млекопитающих
|
|
Оказавшись внутри половых валиков, первичные половые клетки теряют свою подвижность и начинают агрегировать друг с другом. Они продолжают пролиферировать. В это время они называются оогонии и формируют цисты: кластер из взаимосвязанных клеток, полученных в результате митотических делений с неполным цитокинезом (Lei and Spradling, 2013). Цисты также взаимодействуют с соматическими клетками: вместе с эпителиальными прегранулезными клетками они формируют овариальные шнуры, которые отделены от мезенхимных клеток развивающегося яичника базальной мембраной (Pepling, 2006). Оогонии и прегранулезные клетки содержат липидные капли и элементы гладкой эндоплазматической сети, т.е. имеют характеристики стероид-продуцирующих клеток. Под действием ретиноевой кислоты, секретируемой клетками мезонефроса, большинство оогониев вступают в мейоз, с этого момента их называют ооцитами (Smith et al., 2014). У подавляющего большинства млекопитающих на стадии диплотены профазы I наступает блок мейоза (Pepling, 2006).
После рождения морфология яичников быстро изменяется. Происходит реорганизация яичника на функциональные зоны: корковое и мозговое вещество (Wilhelm еt al., 2007). Широкое корковое вещество состоит, главным образом, из фолликулов. Мозговое вещество занимает меньший объем, содержит кровеносные сосуды и гилусные клетки (расположены в области ворот яичника, продуцируют небольшое количество андрогенов). Снаружи яичник покрыт видоизмененным мезотелием - один слой кубических клеток, несущих микроворсинки на апикальных поверхностях. Под эпителием лежит соединительнотканная (белочная) оболочка. Каркас органа представлен веретеновидными фибробластами и фиброцитами (Быков, 1997).
Функциональная единица яичника – фолликул яичника, в котором ооциты созревают в окружении клеток гранулезы и теки. Фолликулы начинают дифференцироваться только после рождения (Wilhelm еt al., 2007).
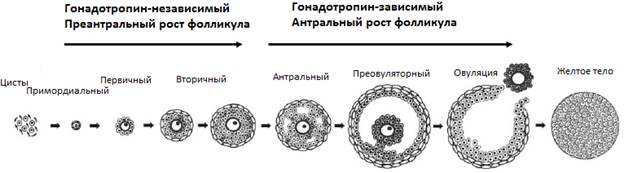
Рис.1. Основные стадии фолликулогенеза у млекопитающих (Edson et al, 2009).
После рождения цисты распадаются на отдельные ооциты. В ходе этого процесса некоторые клетки в каждой цисте уходят в апоптоз, только одна треть от общего числа половых клеток выживает и может быть включена в состав примордиальных фолликулов, каждый из которых состоит из одного ооцита и нескольких плотно прилежащих клеток гранулезы и окружен базальной мембраной (рис. 2). (Pepling, 2006). Клетки гранулезы соединены между собой и с ооцитом многочисленными щелевыми контактами (Сапин и Билич, 2007). Благодаря наличию нексусов клетки гранулезы обеспечивают ооцит питательными веществами (Buccione et al., 1990).
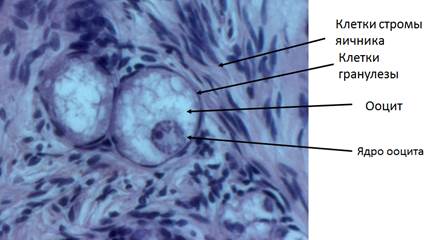
Рис. 2. Примордиальные фолликулы кошки. Объектив 40х.
При формировании первичных фолликулов ооцит увеличивается в размере и производит zona pellucida. Клетки гранулезы из уплощенных становятся кубическими (рис. 3) (Wilhelm еt al., 2007).
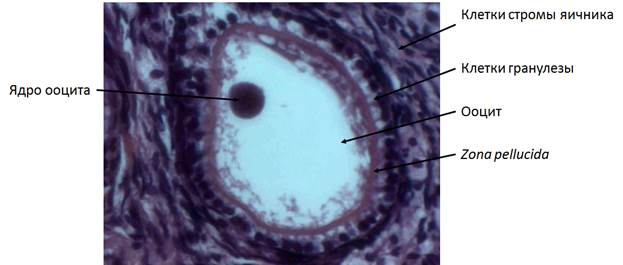
Рис. 3. Первичный фолликул кошки. Объектив 40х.
Формирование вторичного фолликула (рис. 4) характеризуется ростом ооцита, пролиферацией клеток гранулезы до двух и более слоев (Edson et al, 2009). В это время фолликул мигрирует в более васкуляризованную область яичника – мозговое вещество и приобретает дополнительный слой соматических клеток - теку (Futterweit, 1984). Клетки теки дифференцируются из мезенхимных клеток стромы яичника. Тека имеет две зоны: theca interna и theca externa. Клетки theca interna, которые образуются в непосредственной близости от базальной мембраны, окружающей клетки гранулезы, имеют многочисленные митохондрии с трубчатыми кристами, хорошо развитый гладкий эндоплазматический ретикулум и липидные капли – все это соответствует их основной функции – продукция андрогенов для клеток гранулезы, которые преобразуют их в эстрогены (т.к. они содержат ароматазу) (Edson et al, 2009).
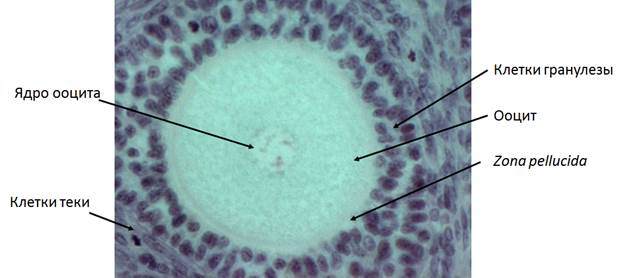
Рис. 4. Вторичный фолликул кошки. Объектив 40х.
Преантральный фолликулогенез является гормононезависимым и контролируется внутригонадными факторами. Развитие фолликула требует двунаправленной связи между яйцеклеткой и клетками гранулезы, клетками гранулезы и клетками теки.
На более поздних стадиях фолликулогенеза (антральный фолликулогенез) контроль роста клеток грунулезы и теки осуществляется внегонадными факторами (Wilhelm еt al., 2007).
На следующем этапе фолликулогенеза формируются несколько маленьких заполненных жидкостью полостей (рис. 5). Они образуются либо в местах с меньшим числом контактов, либо там, где клетки гранулезы умерли. Фолликулярная жидкость, вероятно, происходит из крови, протекающей по капиллярам теки. Накопление жидкости происходит за счет высокого осмотического давления, которое возникает из-за глюкозаминогликанов (гиалуроновой кислоты и хондроитинсульфата), секретируемых клетками гранулезы (Rodgers and Irving-Rodgers, 2010). Полости, заполненные фолликулярной жидкостью, в конечном счете, сливаются, формируя антральную полость, которая разделяет две функционально различные популяции клеток гранулезы. Клетки гранулезы, расположенные вдоль стенки фолликула имеют важнейшее значение для стероидогенеза и овуляции, тогда как клетки кумулюса, окружающие ооцит, способствуют его росту и компетенции развития. Эти два типа клеток зависят от противоположных градиентов ФСГ извне и ооцит-секретируемых факторов изнутри фолликула.
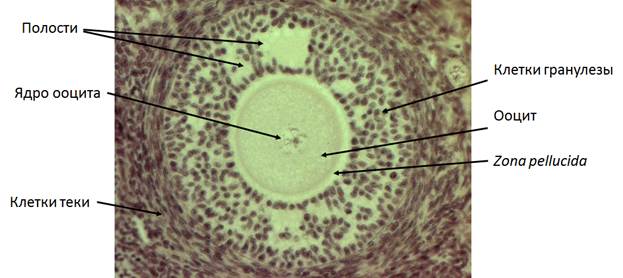
Рис. 5. Антральный фолликул кошки. Объектив 20х.
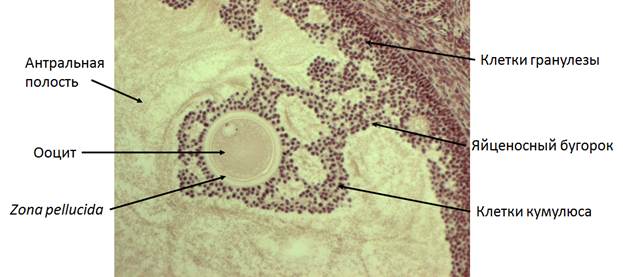
Рис. 6. Фрагмент проевуляторного фолликула кролика. Объектив 10х.
Фолликулы в яичнике созревают не одновременно. В любой момент времени можно найти фолликулы на каждом этапе развития. Тем не менее, не все фолликулы в конечном счете проходят овуляцию успешно. Многие претерпевают атретические изменения (рис. 7) (дегенеративный процесс, включающий гибель клеток гранулезы путем апоптоза и впоследствии утрату ооцита), но один или несколько антральных фолликулов (число зависит от вида) достигают преовуляторного этапа (Wilhelm еt al., 2007).
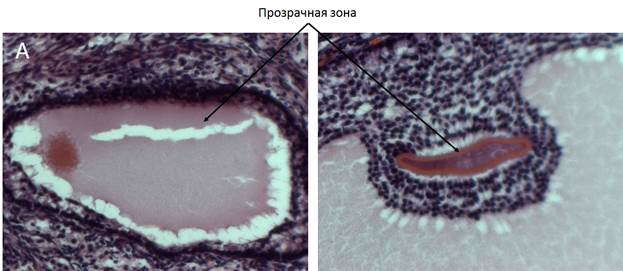
Рис. 7. Атретичекие фолликулы кошки: А – первичный, Б – третичный. Объектив 20х.
Во время овуляции происходит возобновление мейоза в ооците (до метафазы II), разрыв фолликула и выход ооцит-кумулюсного комплекса. Оставшиеся клетки гранулезы и теки формируют желтое тело (рис. 8), высоко дифференцированную эндокринную структуру, секретирующую прогестерон, который подготавливает матку и поддерживает беременность. Процесс формирования желтого тела требует выхода клеток гранулезы из клеточного цикла.
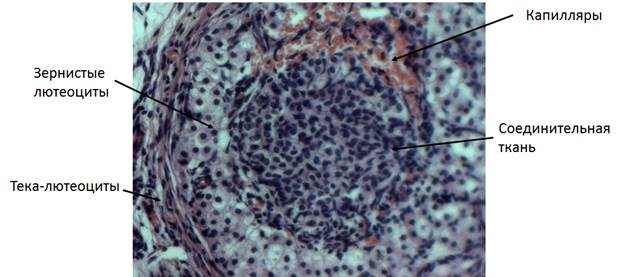
Рис. 8. Желтое тело яичника кошки. Объектив 20х.
Мейоз останавливается на стадии метафазы II из-за активности CSF (цитостатического фактора). CSF стабилизирует MPF (фактор ускорения созревания), который состоит из комплекса CDK1 (циклин зависимой киназы) и циклина В, путем ингибирования APC/C (комплекс стимуляции анафазы), убиквитинлигаза которого связывается с циклином B и вызывает его деградацию и, как следствие, переход клетки к анафазе.
Ооцит проходит финальные стадии мейоза уже после оплодотворения. Вход сперматозоида вызывает волну Ca2+, что активирует APC/C, это приводит к разрушению циклина В и завершению мейоза (Edson et al., 2009).
Список литературы
Быков B.Л. Частная гистология человека (краткий обзорный курс). 2-е изд. – СПб.: СОТИС. – 2002. – 304.
Голиченков В.А., Иванов Е.А., Лучинская Н.Н. и др. Практикум по эмбриологии: Учеб. Пособие для студ. Университетов. – М.: Издательский центр «Академия». - 2004. – 208с.
Сапин М.Р., Билич Г.Л. Анатомия человека: Учеб. для студентов высш. учеб. заведения: В 2 кн. Кн. 2. – 7-е изд., перераб. И доп. – М.: ООО «Издатеьство Оникс»: ООО «Издательство «Мир и Образование», 2007. – 480 с.
Buccione R., Schroeder A.C., Eppig J. J. Interactions between Somatic Cells and Germ Cells throughout Mammalian Oogenesis.// BIOLOGY OF REPRODUCTION. – 1990. – V.43. - P. 543-547.
Edson M.A., Nagaraja A.K., Matzuk M.M. The Mammalian Ovary from Genesis to Revelation.// Endocrine Reviews. – 2009. – V. 30. – P. 624–712.
Futterweit W. Polycystic Ovarian Disease. Springer-Verlag New York Inc – 1984.
Lei L., Spradling A.C. Mouse primordial germ cells produce cysts that partially fragment prior to meiosis.// Development. – 2013. – V. 140. – P. 2075-2081.
Pepling M.Е. From Primordial Germ Cell to Primordial Follicle: Mammalian Female Germ Cell Development.//genesis. – 2006. – V. 44. – P. 622 – 632.
Rodgers R. J., Irving-Rodgers H. F. Formation of the Ovarian Follicular Antrum and Follicular Fluid.// BIOLOGY OF REPRODUCTION. – 2010. – V.82. – P.1021–1029.
Smith P., Wilhelm D., Rodgers R.J. Development of mammalian ovary.// Society for Endocrinology. – 2014. – V.221: 3. – P.145-161.
Wilhelm D., Palmer S., Koopman P. Sex Determination and Gonadal Development in Mammals.// Physiol Rev. – 2007. – V. 87. – P. 1–28.