Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Это единый природный комплекс, который выступает как функциональное целое и образован живыми организмами и средой обитания.
|
|
Основными свойствами экосистемы являются:
- Способность производить органическую продукцию, при этом связывая и высвобождая энергию.
- Способность осуществлять круговорот веществ.
- Способность противостоять внешним воздействиям.
Блоковая модель экосистемы. Любая экосистема состоит из двух блоков. Один из них представлен комплексом взаимосвязанных живых организмов – биоценозом, а второй – факторами среды – биотопом (экотопом). Т.о., в экосистему (биогеоценоз) входит две составляющие: биоценоз и биотоп. В.Н.Сукачев блоковую модель биогеоценоза представлял следующей схемой.
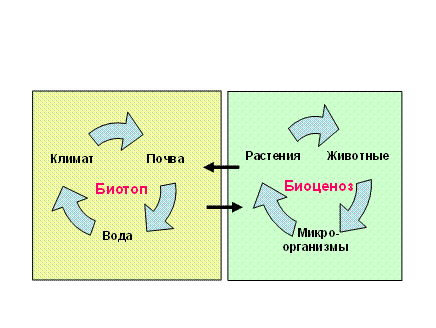
Схема биогеоценоза (экосистемы) по В.Н.Сукачеву
Трофическая структура экосистем. Цепи питания.Любая экосистема включает несколько трофических уровней. Первый уровень представлен растениями. Их называют продуцентами из-за способности продуцировать органическое вещество в процессе фотосинтеза. Второй уровень представлен животными организмами. Они являются гетеротрофами по способу питания и их пищей служат растения. Этот уровень называют консументами первого порядка.Третий уровень (также как и четвертый, пятый) представлен хищниками или консументами второго (или соответствующего) порядка. Один и тот же организм может в различных цепях питания занимать разные трофические уровни, являясь в одном случае консументом 1-го, а в другом – 2-го или 3-го порядка. Это связано с тем, что большинство животных являются эврифагами, т.е. их рацион может быть достаточно разнообразным. Последний уровень представлен организмами, питающимися мертвым органическим веществом: микроорганизами, грибами, мелкими животными. Их называют редуцентами.
Взаимосвязанный ряд трофических уровней представляет цепь питания или трофическую цепь. Главное свойство цепи питания – осуществление биологического круговорота веществ и высвобождение запасенной в органическом веществе энергии.
Цепи питания не всегда могут быть полными. В них могут отсутствовать растения. Такая цепь питания характерна для сообществ, формирующихся на базе разложения трупов животных или растительных остатков и называют ее детритной или цепью разложения. Схематично можно представить ее следующим образом:
Детрит → детритофаги → хищники детритофагов
(редуценты) (консументы 2, 3...пор.)
Цепи питания, которые начинаются с растений, называют пастбищными или цепями выедания:
Растения → травоядные → хищники
(продуценты) (консументы 1пор.) (консументы 2 пор.)
Цепи питания являются тесно взаимосвязанными. Так, например, детрит образуется из продуктов жизнедеятельности всех организмов экосистемы, а также их останков. Кроме того, один и тот же организм может быть участником и детритной и пастбищной цепи. Совокупность всех переплетающихся пищевых цепей экосистемы образует трофическую сеть. Таким образом, цепи питания выделяют искусственно при изучении трофической структуры экосистем.
Исходя из положения: разнообразие – синоним устойчивости, можно заключить, что чем длиннее цепи питания и чем сложнее трофическая структура экосистемы, тем она устойчивее.
Живые организмы, входящие в экосистемы, для своего существования должны постоянно пополнять и расходовать энергию. Растения запасают энергию в процессе фотосинтеза, превращая энергию Солнца в энергию химических связей. При фотосинтезе связывается энергия только с определенной длиной волны – 380-710 нм. Эту энергию называют фотосинтетически активной радиацией (ФАР). Она по длинам волн близка к видимой части спектра. На эту радиацию приходится обычно около 40% общей солнечной радиации, достигающей земной поверхности.
Растения в процессе фотосинтеза связывают лишь небольшую часть солнечной радиации. Даже по отношению к фотосинтетически активной – это в среднем для Земного шара около 1 %. Только наиболее продуктивные экосистемы, такие как плантации сахарного тростника, посевы кукурузы, тропические леса в оптимальных условиях могут связывать до 3-5 % ФАР.
Растения являются первичными поставщиками энергии для всех других организмов в цепях питания. Существуют определенные закономерности перехода энергии с одного трофического уровня на другой вместе с потребляемой пищей. Основная часть энергии, усвоенной консументами с пищей, расходуется на его жизнеобеспечение (движение, поддержание температуры тела и т.п.). Эту часть энергии рассматривают как траты на дыхание, с которым, в конечном счете, связаны все возможности ее высвобождения из химических связей органического вещества. Часть энергии переходит в тело потребителя, увеличивая его биомассу. Некоторая доля пищи не усваивается организмом, а следовательно, из нее не высвобождается и энергия. В последующем она высвобождается из экскрементов, но другими организмами, которые потребляют их в пищу. Особенно велики потери энергии с экскрементами у травоядных животных (у некоторых – до 70 %).
Переход энергии с одного трофического уровня на другой в среднем принимается близким к 10 % от энергии, потребленной с пищей. Эта закономерность рассматривается обычно как «правило 10 %». Из него следует, что цепь питания имеет ограниченное количество уровней, обычно не более 4-5. Пройдя через них, практически вся энергия оказывается рассеянной. Из данной закономерности вытекают важные в практическом отношении практические выводы.
1. С энергетической точки зрения крайне нецелесообразно потребление животной продукции, особенно с высоких уровней цепей питания.
2. Для решения продовольственной проблемы в условиях демографического взрыва надо, чтобы в рационе людей больший удельный вес составляла растительная пища. Энергетически идеально – вегетарианство.
3. Для увеличения КПД использования кормов при получении животноводческой продукции очень важно уменьшить основную статью нерационального расходования энергии – ее траты на дыхание. Это возможно за счет поддержания оптимального температурного режима в животноводческих помещениях, ограничения подвижности животных, сбалансированности кормового рациона по различным элементам питания, а также применения различных добавок (для стимуляции роста, улучшения аппетита и т.д.).
Одно из важнейших свойств организмов, их популяций и экосистем в целом – способность создавать органическое вещество, которое называют продукцией. Образование продукции в единицу времени на единице площади или объема, выраженное в единицах массы, характеризует продуктивность экосистем.
Продукцию растений называют первичной, а животных – вторичной.
Наряду с продукцией различают биомассу организма или экосистемы в целом. Под ней понимают все живое вещество, которое содержится в экосистеме или ее элементах вне зависимости от того, за какой период времени она образовалась.
Величина биомассы экосистем зависит не столько от их продуктивности, сколько от продолжительности жизни организмов и экосистем. Например, большая биомасса характерна для лесных экосистем: в тропических лесах она достигает 800-1000 т/га, в лесах умеренной зоны – 300-400 т/га. Для экосистем, представленных однолетними организмами, их годичная продуктивность и биомасса практически совпадают.
Если количество энергии, продукции, биомассу или численность организмов на каждом трофическом уровне изображать в виде прямоугольников в одном и том же масштабе, то их распределение будет иметь вид пирамид. Эта закономерность носит название правило пирамид. В отношении энергии можно сказать, что количество энергии, содержащейся в организмах на любом последующем трофическом уровне цепи питания, меньше ее значений на предыдущем уровне. 
1 2
Пирамиды продукции, энергии (биомасс для экосистем суши) – 1 и биомасс для экосистем океана – 2.
Количество продукции, образующейся в единицу времени на разных трофических уровнях, подчиняется тому же правилу, которое характерно для энергии: на каждом последующем уровне количество продукции меньше, чем на предыдущем.
Пирамиды биомассы сходны с таковыми для энергии и продукции только для сухопутных экосистем. Для водных экосистем пирамида биомасс как бы перевернута. Это означает, что биомасса животных, потребляющих растительную продукцию, больше биомассы растительных организмов. Причина этого – резкие различия в продолжительности жизни организмов сравниваемых уровней. Первый уровень представлен в основном фитопланктоном с крайне короткой продолжительностью жизни (несколько дней или даже часов), второй и более высокие – более долгоживущими организмами – зоопланктоном, рыбами, моллюсками и т.д. Они накапливают биомассу годами и десятилетиями.
Пирамида чисел свидетельствует, что количество организмов, как правило, уменьшается от основания к вершине. Это правило не абсолютно и применимо в основном к цепям питания, не включающим редуцентов.
Любая экосистема, приспосабливаясь к изменениям внешней среды, находится в состоянии динамики. Эта динамика может касаться как отдельных ее звеньев, так всей экосистемы в целом.
Динамика может быть периодической – суточной, сезонной, а может быть непериодической, направленной, которая называется развитием или сукцессией.
Сукцессия – процесс последовательной смены одной экосистемы другой, преемственно возникающей на том же месте под действием внешних или внутренних факторов. При этом в экосистему могут внедряться новые виды, либо одни виды могут сменяться другими.
Если сукцессия обусловливается внешними факторами, то ее называют экзогенетической, если внутренними – эндогенетической. Примером экзогенетической сукцессии могут служить изменения экосистем под влиянием потепления климата, иссушения почв, понижения уровня грунтовых вод и др. Их называют вековыми сукцессиями, т.к. длиться они могут столетиями и тысячелетиями. В качестве эндогенетической сукцессии рассмотрим изменения заброшенного пахотного участка. В качестве общих закономерностей будет иметь место заселение живыми организмами, увеличение их видового разнообразия, постепенное обогащение почвы органическим веществом, усиление связей между различными видами, уменьшение числа свободных экологических ниш, постепенное формирование все более сложных биоценозов и экосистем, повышение их продуктивности. При этом всегда можно выделить последовательные стадии сукцессий (сукцессионные ряды), которые заканчиваются относительно мало изменяющимися экосистемами, которые называются коренными, узловыми или климаксными. Видовой состав климаксных экосистем может существенно различаться, общим является лишь то, что в каждой из них имеются виды-эдификаторы, которые в наибольшей мере создают среду обитания. Например, для степных экосистем эдификаторами являются плотнокустовые злаки; для тропических лесов это древесные виды, создающие сильное затенение для других видов.
Различают следующие виды сукцессий:
1. первичные – они начинаются с исходно безжизненного субстрата, примером которого могут служить песчаные обнажения, горные отвалы, застывшая после извержения вулкана лава;
2. вторичные – они возникают на месте нарушенных или разрушенных экосистем, например, после вырубки леса, лесных пожаров, при зарастании площадей, находившихся ранее под сельскохозяйственными угодьями.
Вторичные сукцессии протекают несравненно быстрее первичных, т.к. начинаются с промежуточных стадий и на фоне более богатых почв.
Различают также автотрофные и гетеротрофные сукцессии. Автотрофные протекают в экосистемах, где центральным звеном является растительный покров. Такие сукцессии потенциально бессмертны, поскольку все время пополняются энергией и веществом в процессе фотосинтеза. К гетеротрофным относятся те сукцессии, которые протекают в субстратах, где отсутствуют живые растения, а участвуют только животные или мертвые растения. Этот вид сукцессий имеет место только до тех пор, пока присутствует запас готового органического вещества. Эта сукцессия по своей природе деструктивна. Примером такой сукцессии может служить разложение мертвого дерева или трупа животного.
Общие закономерности сукцессионного процесса.
1. На начальных стадиях видовое разнообразие, продуктивность и биомасса малы. По мере развития сукцессии эти показатели возрастают.
2. С развитием сукцессионного ряда увеличиваются взаимосвязи между организмами. Полнее осваивается среда обитания, усложняются цепи и сети питания.
3. Уменьшается количество свободных экологических ниш и в климаксном сообществе они либо отсутствуют, либо находятся в минимуме.
4. Интенсифицируются процессы круговорота веществ, потока энергии и дыхания экосистем.
5. Неизменяемость климаксных стадий сукцессий относительна. Динамические процессы при этом не приостанавливаются, а лишь замедляются.
6. В зрелой стадии климаксного сообщества биомасса обычно достигает максимальных или близких к максимальным значений.
Стабильность – способность экосистемы сохранять свою структуру и функциональные свойства при воздействии внешних факторов.
Устойчивость – способность экосистемы возвращаться в исходное или близкое к нему состояние после действия факторов, выводящих ее из равновесия.
Данные термины в экологии обычно рассматриваются как синонимы. Эти качества тем значительнее, чем разнообразнее экосистемы.
Для экосистем с низкой устойчивостью характерны вспышки отдельных видов. Низкоустойчивыми и нестабильными являются, например, тундровые экосистемы, которые легко разрушаются под действием перевыпаса, технических нагрузок. К таким уязвимым экосистемам относятся также агроэкосистемы, созданные человеком и представленные обычно одним преобладающим видом растений, интересующим человека. Агроценозы сельскохозяйственных культур, особенно однолетних, существуют только при условии постоянного вмешательства человека. После прекращения такого вмешательства вторичная сукцессия обычно начинается с той стадии, которую называют сорняками. Но эта стадия уже не имеет прямого отношения к агроценозу. Агроценозы, создаваемые из долгоживущих растений (искусственно созданные лесопосадки, сады), отличаются значительной устойчивостью. Здесь вмешательство человека требуется только на начальных этапах, когда молодые деревца настолько слабы, что не могут выдержать конкуренции с травами.