Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Наброски новой концепции
|
|
Как показано в предпринятом выше анализе, гипертрофия и атрофия скелетных мышц в общем случае могут быть следствием как изменения интенсивности транскрипции мРНК в ядрах мышечных клеток, так и следствием изменения числа ядер в мышце — но при этом итоговый вклад данных факторов в результат двух антагонистичных адаптационных процессов весьма различен.
При развитии функциональной гипертрофии мышц доминирует следующая последовательность событий:
Увеличение нагрузки на мышцы —> активация пролиферации миосателлитоцитов —> увеличение числа ядер в мышце —> синтез РНК на новых ядрах —> синтез новых сократительных структур —> гипертрофия мышц
Снижение двигательной активности мышц, в свою очередь, активизирует вот какую последовательность событий, приводящих к атрофии мышц:
Снижение двигательной активности мышц —> снижение интенсивности транскрипции мРНК структурных белков и снижение пролиферативной активности миосателлитоцитов —> уменьшение размера ДНК-единицы и снижение числа ядер по мере их апоптоза —> атрофия мышц
Ввиду ограниченности размера ДНК-единицы изменение интенсивности транскрипции мРНК структурных белков играет важную роль в процессах атрофии мышц, но не в процессах их гипертрофии. Вместе с тем следует отметить, что от интенсивности транскрипции мРНК структурных белков зависит не только размер ДНК-единицы. Посредством управления интенсивностью экспрессии генов осуществляется регуляция спектра синтезируемых белков, что оказывает кардинальное влияние на функциональные свойства мышц.
Сравнение состава мышц крыс после компенсаторной гипертрофии, вызванной отсечением мышц-синергистов, и после функциональной гипертрофии, вызванной регулярным принудительным плаванием, показало, что компенсаторная гипертрофия сопровождается увеличением плотности митохондрий, уменьшением плотности миофибрилл и неизменностью плотности саркоплазматического ретикулума. В свою очередь, функциональная гипертрофия сопровождается увеличением плотности саркоплазматического ретикулума, а плотность митохондрий и миофибрилл остаётся неизменной (D.Seiden 1976).
В результате тренировок в мышцах может возрастать концентрация одних ферментов, обеспечивающих воспроизводство энергии, при неизменности концентрации других ферментов — в результате чего мышцы изменяют свои окислительные или гликолитические возможности (N.Wang et al. 1993).
Под воздействием тренировки возможно изменение характерных свойств мышечных волокон вплоть до изменения типа волокон (F.Ingjer 1979), (R.S.Staron et al. 1990), (N.Wang et al. 1993).
Изменения структуры и свойств мышц под воздействием тренировки не исчерпываются приведёнными выше примерами, но рассмотрение этих изменений не является темой настоящего исследования. Данные примеры были приведены лишь для того, чтобы показать, что изменения, происходящие с мышечными волокнами в результате тренировки, могут быть связаны с изменением белкового состава волокон, то есть могут являться следствием изменения интенсивности транскрипции мРНК различных видов структурных белков. Соответственно, воздействие тренировки на генетический аппарат мышечной клетки не может сводиться к усилению общего синтеза белка через общий для всех структурных белков фактор-регулятор. Более того, интенсификация синтеза некоторых видов сократительных белков происходит отнюдь не только при возрастании двигательной активности мышц. Так, снижение нагрузки на мышцы крыс, вызванное пребыванием животных в невесомости, снижает в ряде медленных волокон синтез цепей миозина, характерных для медленных волокон, но увеличивает экспрессию некоторых форм быстрого миозина (D.L.Allen et al 1996). Обратным образом, функциональная перегрузка мышц кошек снижает экспрессию некоторых форм быстрого миозина в медленных волокнах (D.L.Allen et al 1995). Эти факты не укладываются в концепцию прямого активирующего воздействия факторов энергетического истощения на экспрессию мРНК сократительных белков. Экспрессия мРНК сократительных белков мышц если и зависит от метаболических факторов, то зависимость эта проявляется, по-видимому, более сложным образом.
Как было отмечено в начале данного текста, часть спортивных исследователей отводит роль регулятора транскрипции мРНК сократительных белков мышц креатину, однако роль креатина в регуляции синтеза сократительных белков нельзя считать однозначно установленной. Действительно, в ряде исследований (J.S.Ingwall et al. 1972), (J.S.Ingwall et al. 1974), (M.L.Zilber et al. 1976) было показано, что повышение концентрации креатина интенсифицирует синтез специфических мышечных белков (миозина и актина) в развивающихся мышечных клетках in vitro. Эти наблюдения были восприняты как важное свидетельство того, что индуктором транскрипции сократительных белков является именно креатин. Однако впоследствии в противовес упомянутым выше исследованиям влияние креатина на синтез миозина обнаружено не было (D.M.Fry, M.F.Morales 1980), (R.B.Young, R.M.Denome 1984). R.B.Young и R.M.Denome предположили, что уровень креатина может регулировать синтез миозина лишь на ранних этапах эмбрионального развития мышечных клеток, но не может являться регулятором синтеза сократительных белков в уже сформированных мышечных волокнах.
Таким образом, гипотеза о роли креатина в регуляции синтеза сократительных белков требует дальнейшей проверки. Тем не менее, исходя из общих соображений, следует признать, что концепция, согласно которой индуктором транскрипции мРНК структурных белков является креатин или какой-либо иной фактор, связанный с истощением энергетики мышц, кажется достаточно убедительной лишь в отношении регуляции синтеза мышечных ферментов — если предположить, что регуляция ферментного синтеза в сложных многоклеточных организмах осуществляется по тому же принципу, что и у прокариот. Такие метаболилы, как АДФ, АМФ, ортофосфат, креатин и др., накапливающиеся в активно сокращающихся мышечных волокнах, сами являются субстратами для реакций, восстанавливающих запас энергетических фосфатов в волокне. Соответственно, накопление в мышцах указанных метаболилов должно стимулировать транскрипцию мРНК ферментов, которые обеспечивают протекание энерговосстанавливающих реакций, использующих данные метаболилы в качестве субстратов. Регулярная работа до утомления мышц должна сопровождаться регулярной активацией синтеза ферментов и в конечном итоге приводить к их накоплению в мышцах. Обратным образом, снижение двигательной активности мышц должно снижать частоту активирования синтеза мРНК ферментов. Соответственно, содержание в мышцах ферментов по мере естественного катаболизма последних должно убывать. Предположение, что накопление в мышцах ферментов происходит вследствие субстратно-индуцируемого усиления синтеза данных ферментов, выдвинул ещё Н.Н.Яковлев (Н.Н.Яковлев 1974). Ф.З.Меерсон в обоснование гипотезы о влиянии ацидоза мышц на индукцию мРНК структурных белков привёл аргументы, которые также касаются индукции синтеза белков, ответственных именно за энергообеспечение мышц. Меерсон отметил, что ацидоз мышц является ранним сигналом энергетической недостаточности, и потому с позиций эволюционной теории оправданно будет предположить, что этот же сигнал вполне может использоваться в качестве активатора генетического аппарата клетки. В конечном итоге сие должно приводит к росту структур, которые призваны устранять дефицит энергии — и организм тем самым становится в целом более резистентными к изменившимся условиям внешней среды (Ф.З.Меерсон 1993).
Такую аргументацию можно признать весьма убедительной, но вот расширение данного принципа на регулирование синтеза иных видов мышечных белков, в особенности сократительных (как это происходит в концепции того же Меерсона и многих иных исследователей), кажется уже не совсем оправданным именно с эволюционной точки зрения. Высокая концентрация в саркоплазме продуктов распада макроэнергетических фосфатов является сигналом о том, что возможности мышечного волокна в восстановлении уровня АТФ за счёт окислительных процессов и гликолиза недостаточны для данной интенсивности сокращения. В такой ситуации адаптация мышечного волокна должна быть направлена в сторону увеличения мощности восстанавливающих энергию реакций. Синтез же сократительных белков (основных потребителей энергии) может способствовать лишь увеличению скорости расхода АТФ в волокне и приводить к ещё большему падению уровня АТФ при новых аналогичных нагрузках — поэтому адаптация в данном направлении не может сделать мышечные волокна более устойчивыми к изменившейся требованиям в отношении двигательной активности мышц.
Таким образом, стимулы к развитию энергетики мышц и стимулы к экстенсивному развитию сократительного аппарата мышц должны иметь и, похоже, имеют разную природу.
Как уже было отмечено выше, совершенствование энергетических возможностей мышц тесно связано с увеличением содержания в мышцах ферментов, то есть является следствием субстратно-индуцируемого активирования транскрипции мРНК данных видов белка. Вполне вероятно, что синтез мРНК иных видов белка, связанных с энергообеспечением мышц (например, миоглобина или митохондриальных белков) может происходить по схожей схеме. Но, как показано выше, размер ДНК-единицы ограничен и каждое клеточное ядро ответственно за поддержание функционирования строго определённого объёма мышечного волокна. Для кардинального увеличения объёма мышц и строительства в них новых сократительных структур нужны новые клеточные ядра в дополнение к уже существующим, то есть экстенсивное развитие мышц связано в первую очередь с активацией пролиферации клеток-сателлитов. Вместе с тем очевидно, что поскольку белковый состав сократительных структур различен для различных типов волокон и зависит от режима функционирования мышц, сигналы какого-то иного рода, воздействующие на генетический аппарат мышечных клеток, должны дополнительно регулировать спектр экспрессируемых сократительных белков.
Приведённый в настоящем тексте анализ показал, что общепринятая схема взаимосвязи звеньев срочной и долговременной адаптации мышц к нагрузке (см. рис. 2)
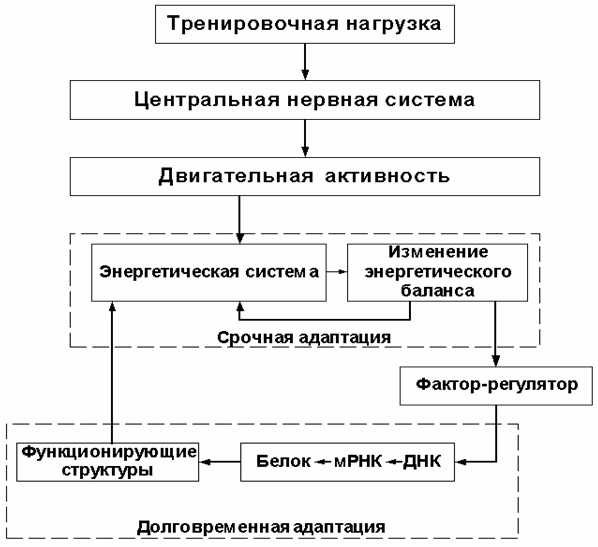
Рисунок 2
в отношении скелетных мышц описывает лишь часть адаптационных процессов, а именно — адаптацию энергетической системы мышц. Эта схема оставляет без внимания ряд важных механизмов долговременной адаптации скелетных мышц к нагрузке, а потому требует значительного уточнения (см. рис. 3).
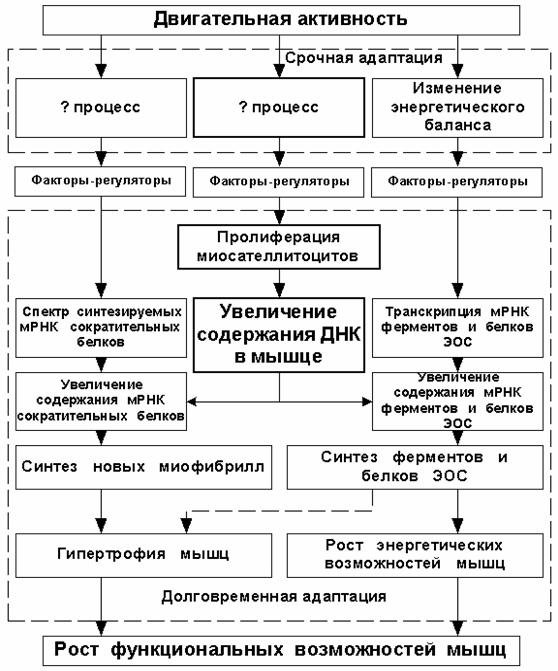
Рисунок 3 (ЭОС — энергообеспечивающие системы)
Следует отметить, что предлагаемая блок-схема механизмов адаптации мышц к нагрузке также не является исчерпывающей, поскольку не включает в себя достаточно важные механизмы гормональной адаптации организма к нагрузке — в ней учтены лишь основные локальные (внутримышечные) адаптационные процессы, которые только и были предметом рассмотрения настоящего исследования.
Возникает вопрос: каковы последствия такого изменения принципиальной схемы адаптации для теории спортивной тренировки, то есть имеет ли для разработки тренировочных методик и планирования нагрузки значение то, за счёт каких именно процессов происходит адаптация мышц? Ответ на данный вопрос таков: да, судя по всему, изменение представлений о схеме адаптации мышц к нагрузке имеет немалое значение.
Дело в том, что интенсивная сократительная активность мышц блокирует синтез белка в мышцах и даже активизирует его катаболизм. Следовательно, рациональным следует считать такой режим тренировок, при котором новое тренировочное занятие сочетается по времени с отключением адаптивного синтеза белка после предыдущего тренировочного занятия или же со значительным снижением его интенсивности (А.А.Виру, Н.Н.Яковлев 1988). Если при реализации данного принципа исходить из того, что тренировочное воздействие сводится лишь к активированию транскрипции мРНК структурных белков под воздействием единого фактора-регулятора, то максимальный эффект будет иметь место в результате применения крайне простой тренировочной схемы со следующими друг за другом через равные интервалы отдыха тренировочными занятиями, интенсивность которых возрастает по мере тренированности организма. Однако, к сожалению, из спортивной практики хорошо известна малая эффективность такого рода тренировочных методик, в особенности для хорошо тренированных спортсменов.
Из предложенной в настоящем тексте схемы развития долговременной адаптации скелетных мышц (см. рис. 3) видно, что адаптивное усиление синтеза белка связано не только с процессами активирования транскрипции мРНК структурных белков, но и с увеличением объёма синтезированного белка за счёт синтеза белка на мРНК, экспрессируемых новообразованными ДНК. Причём посттренировочное активирование транскрипции мРНК играет наиболее важную роль в регуляции синтеза белков, связанных с энергообеспечением мышц. Для повышения энергетических возможностей мышц тренировочные занятия, активирующие транскрипцию мРНК белков энергообеспечивающих систем, следует проводить в такой период, когда адаптивный синтез данных белков, вызванный предыдущим тренировочным занятием, близок к завершению или, во всяком случае, прошёл фазу наивысшей активности.
Адаптивное же усиление синтеза белка за счёт мРНК, экспрессируемых новообразованными ядрами, можно считать завершённым только тогда, когда завершено строительство новых сократительных структур на основе новообразованных ядер, то есть когда в мышцах после увеличения числа ядер восстановлен характерный размер ДНК-единицы. Строительство сократительных структур с нуля в отличие от синтеза ферментов — процесс весьма длительный, поэтому оптимальная частота тренировочных занятий, активирующих пролиферацию миосателлитоцитов, может кардинально отличаться от оптимальной частоты тренировок, обеспечивающей максимальный синтез белков энергообеспечивающих систем мышц.
В предлагаемой блок-схеме локальных механизмов долговременной адаптации скелетных мышц два блока отмечены знаком вопроса, а факторы-регуляторы не определены. Как уже отмечалось выше, факторами-регуляторами синтеза ферментов являются продукты энергетического метаболизма, а вот совокупность факторов, влияющих на спектр экспрессируемых сократительных белков, а также факторы, активирующие пролиферацию миосателлитоцитов, до настоящего времени в полной мере не установлены. Продвижение исследований в данных направлениях позволит в будущем разработать более специализированные методы тренировки, направленно стимулирующие различные адаптационные процессы в мышцах. В свою очередь, более чёткое разделение тренировочного воздействия позволит оптимизировать дозирование нагрузки в тренировочном микроцикле.
Литература:
1. Аракелян Е.Е., Збарский В.А., Шевченко Т.Н., Селуянов В.Н." Формирование техники спринтерского бега посредством направленного развития силы отдельных мышечных групп у юных атлетов" Физическая культура: воспитание, образование, тренировка. 1997 г. N 3
2. Березов Т.Т., Коровкин Б.Ф. " Биологическая химия" Изд. " Медицина" Москва 1998 г. 704 стр.
3. Балаболкин М.И. " Эндокринология" Москва " Универсум паблишинг" 1998 г.
4. Виру А.А., Яковлев Н.Н. " Главы из спортивной физиологии" Изд. Тартуского государственного университета. Тарту 1988 г. 134 стр.
5. Волков Н.И., Нессен Э.Н., Осипенко А.А., Корсун С.Н. " Биохимия мышечной деятельности" Изд. " Олимпийская литература" Киев 2000 г. 503 стр.
6. Володина А.В. " Посттравматическая регенерация скелетных мышц" Диссертация доктора биологических наук. Москва 1995 г.
7. Данилов Р.К. " Очерки гистологии мышечных тканей" Уфа 1994 г. 49 стр.
8. Калинский М.И., Курский М.Д., Осипенко А.А. " Биохимические механизмы адаптации при мышечной деятельности" Киев. Изд. " Выща школа" 1986 г. 183 стр.
9. Мак-Комас А.Дж. " Скелетные мышцы" Изд. " Олимпийская литература" Киев 2001 г. 407 стр.
10. Меерсон Ф.З " Адаптационная медицина: концепция долговременной адаптации" Москва. Изд. " Дело" 1993 г. 138 стр.
11. Меерсон Ф.З. Пшенникова М.Г. " Адаптация к стрессовым ситуациям и физическим нагрузкам" Москва. Изд. " Медицина" 1988 г. 253 стр.
12. Селуянов В.Н. " Разработка методов и планов физической подготовки спортсменов на основе имитационного моделирования" Из сборника " Теория и практика применения дидактики развивающего обучения в подготовке специалистов по физическому воспитанию" Изд. " Физкультура, образование и наука". Труды сотрудников проблемной научно-исследовательской лаборатории / Научный руководитель В.Н.Селуянов. — М: Физкультура, образование и наука, 1996. — 106 стр.
13. Сингер М., Берг П. " Гены и геномы" в 2-х томах Изд. " Мир" Москва 1998г. 764 стр.
14. Улумбеков Э.Г., Челышев Ю.А. " Гистология, введение в патологию", Москва " ГЕОТАР Медицина" 1998 г.
15. Шекман Б.С. " Влияние тренировки на композицию мышц, размер и окислительный потенциал мышечных волокон у человека" Диссертационная работа. Москва 1990 г.
16. Шубникова Е.А., Юрина Н.А., Гусев Н.Б., Балезина О.П., Большакова Г.Б. " Мышечные ткани" Москва " Медицина" 2001 г.
17. Яковлев Н.Н. " Биохимия спорта" Изд. " Физкультура и спорт". Москва 1974 г. 288 стр.
18. Яковлев Н.Н., Коробков А.В., Янанис С.В. " Физиологические и биохимические основы теории и методики спортивной тренировки" Изд. " Физкультура и спорт". Москва 1957 г. 323 стр.
19. Allen DL, Linderman JK, Roy RR, Bigbee AJ, Grindeland RE, Mukku V, Edgerton VR (1997) " Apoptosis: a mechanism contributing to remodeling of skeletal muscle in response to hindlimb unweighting" Am J Physiol. 1997 Aug; 273(2 Pt 1): C579-87.
20. Allen DL, Monke SR, Talmadge RJ, Roy RR, Edgerton VR (1995) " Plasticity of myonuclear number in hypertrophied and atrophied mammalian skeletal muscle fibers" J Appl Physiol 78: 1969-1976, 1995;
21. Allen DL, Yasui W, Tanaka T, Ohira Y, Nagaoka S, Sekiguchi C, Hinds WE, Roy RR, Edgerton VR (1996) " Myonuclear number and myosin heavy chain expression in rat soleus single muscle fibers after spaceflight" J Appl Physiol. 1996 Jul; 81(1): 145-51.
22. Allen RE, Rankin LL (1990) " Regulation of satellite cells during skeletal muscle growth and development" Proc Soc Exp Biol Med. 1990 Jun; 194(2): 81-6.
23. Alway SE, Grumbt WH, Gonyea WJ, Stray-Gundersen J (1989a) " Contrasts in muscle and myofibers of elite male and female bodybuilders" J Appl Physiol. 1989 Jul; 67(1): 24-31
24. Alway SE, Winchester PK, Davis ME, Gonyea WJ (1989b) " Regionalized adaptations and muscle fiber proliferation in stretch-induced enlargement" J Appl Physiol. 1989 Feb; 66(2): 771-81.
25. Antonio J, Gonyea WJ (1993 a) " Role of muscle fiber hypertrophy and hyperplasia in intermittently stretched avian muscle" J Appl Physiol. 1993 Apr; 74(4): 1893-8
26. Antonio J, Gonyea WJ (1993 b) " Progressive stretch overload of skeletal muscle results in hypertrophy before hyperplasia" J Appl Physiol. 1993 Sep; 75(3): 1263-71.
27. Appell HJ, Forsberg S, Hollmann W (1988) " Satellite cell activation in human skeletal muscle after training: evidence for muscle fiber neoformation" Int J Sports Med. 1988 Aug; 9(4): 297-9.
28. Bruusgaard JC, Liestol K, Ekmark M, Kollstad K, Gundersen K (2003) " Number and spatial distribution of nuclei in the muscle fibres of normal mice studied in vivo" J Physiol. 2003 Sep 1; 551(Pt 2): 467-78. Epub 2003 Jun 17.
29. Burleigh IG (1977) " Observations on the number of nuclei within the fibres of some red and white muscles" J Cell Sci. 1977 Feb; 23: 269-84.
30. Cabric M, Appell HJ, Resic A (1987) " Effects of electrical stimulation of different frequencies on the myonuclei and fiber size in human muscle" Int J Sports Med. 1987 Oct; 8(5): 323-6.
31. Cabric M, James NT (1983) " Morphometric analyses on the muscles of exercise trained and untrained dogs" Am J Anat.166(3): 359-68.
32. Cheek DB (1985) " The control of cell mass and replication. The DNA unit — a personal 20-year study" Early Hum Dev. 1985 Dec; 12(3): 211-39.
33. Darr KC, Schultz E (1987) " Exercise-induced satellite cell activation in growing and mature skeletal muscle" J Appl Physiol. 1987 Nov; 63(5): 1816-21
34. Dmitrieva EV (1975) " The role of the buds and myoblasts in reparative regeneration of muscle fibers of the skeletal type" Arkh Anat Gistol Embriol. 1975 Feb; 68(2): 37-43.
35. Enesco M, Puddy D " Increase in the number of nuclei and weight in skeletal muscle of rats of various ages" (1964) Amer. J. Anat.114: 235
36. Fry DM, Morales MF (1980) " A reexamination of the effects of creatine on muscle protein synthesis in tissue culture" J Cell Biol. 1980 Feb; 84(2): 294-7.
37. Giddings CJ, Gonyea WJ (1992) " Morphological observations supporting muscle fiber hyperplasia following weight-lifting exercise in cats" Anat Rec.1992 Jun; 233(2): 178-95
38. Gollnick PD, Timson BF, Moore RL, Riedy M (1981) " Muscular enlargement and number of fibers in skeletal muscles of rats" J Appl Physiol. 1981 May; 50(5): 936-43.
39. Gollnick PD, Parsons D, Riedy M, Moore RL (1983) " Fiber number and size in overloaded chicken anterior latissimus dorsi muscle" J Appl Physiol. 1983 May; 54(5): 1292-7.
40. Gonyea W, Ericson GC, Bonde-Petersen F " Skeletal muscle fiber splitting induced by weight-lifting exercise in cats" Acta Physiol Scand. 1977 Jan; 99(1): 105-9.
41. Gonyea WJ, Sale DG, Gonyea FB, Mikesky A (1986) " Exercise induced increases in muscle fiber number" Eur J Appl Physiol Occup Physiol. 1986; 55(2): 137-41.
42. Gustafsson JA, Saartok T, Dahlberg E, Snochowski M, Haggmark T, Eriksson E (1984) " Studies on steroid receptors in human and rabbit skeletal muscle — clues to the understanding of the mechanism of action of anabolic steroid" Prog Clin Biol Res. 1984; 142: 261-90
43. Ingjer F (1979) " Effects of endurance training on muscle fibre ATP-ase activity, capillary supply and mitochondrial content in man" J Physiol. 1979 Sep; 294: 419-32.
44. Ingwall JS, Morales MF, Stockdale FE (1972) " Creatine and the control of myosin synthesis in differentiating skeletal muscle" Proc Natl Acad Sci USA. 1972 Aug; 69(8): 2250-3.
45. Ingwall JS, Weiner CD, Morales MF, Davis E, Stockdale FE(1974) " Specificity of creatine in the control of muscle protein synthesis" J Cell Biol. 1974 Jul; 62(1): 145-51
46. James NT, Cabric M (1981) " Quantitative studies on the numerical frequency of myonuclei in the muscles of exercised rats: evidence against the occurrence of fibre-splitting" Br J Exp Pathol. 1981 Dec; 62(6): 600-5.
47. Kadi F, Eriksson A, Holmner S, Butler-Browne GS, Thornell LE (1999а) " Cellular adaptation of the trapezius muscle in strength-trained athletes" Histochem Cell Biol. 1999 Mar; 111(3): 189-95.
48. Kadi F, Eriksson A, Holmner S, Thornell LE (1999b) " Effects of anabolic steroids on the muscle cells of strength-trained athletes" Med Sci Sports Exerc. 1999 Nov; 31(11): 1528-34.
49. Kasper CE, Xun L (1996) " Cytoplasm-to-myonucleus ratios in plantaris and soleus muscle fibres following hindlimb suspension" J Muscle Res Cell Motil. 1996 Oct; 17(5): 603-10
50. Kelly FJ, Lewis SE, Anderson P, Goldspink DF (1984) " Pre- and postnatal growth and protein turnover in four muscles of the rat" Muscle Nerve. 1984 Mar-Apr; 7(3): 235-42.
51. Kennedy JM, Eisenberg BR, Reid SK, Sweeney LJ, Zak R (1988) " Nascent muscle fiber appearance in overloaded chicken slow-tonic muscle" Am J Anat. 1988 Feb; 181(2): 203-15.
52. Kondalenko VF, Sergeev IP, Ivanitskaia VV (1981) " Electron microscopic study of signs of skeletal muscle fiber hyperplasia in athletes" Arkh Anat Gistol Embriol. 1981 Jun; 80(6): 66-70.
53. Larsson L., Tesch PA (1986) " Motor unit fiber density in extremely hypertriphied skeletal muscles in man" Eur J Appl Physiol Occup Physiol. 1986; 55(2): 130-6.
54. MacDougall JD, Sale DG, Elder GC, Sutton JR (1982) " Muscle ultrastructural characteristics of elite powerlifters and bodybuilders" Eur J Appl Physiol Occup Physiol. 1982; 48(1): 117-26.
55. MacDougall JD, Sale DG, Alway SE, Sutton JR (1984) " Muscle fiber number in biceps brachii in bodybuilders and control subjects" J Appl Physiol. 1984 Nov; 57(5): 1399-403.
56. Manta P, Vassilopoulos D, Spengos M (1987) " Nucleo-cytoplasmic ratio in ageing skeletal muscle" Eur Arch Psychiatry Neurol Sci. 1987; 236(4): 235-6.
57. Mauro A (1961) " Satellite cell of skeletal muscle fibers" J Biophys Biochem Cytol 9: 493-498, 1961
58. McCall, GE, Allen DL, Linderman JK, Grindeland RE, Roy RR, Mukku VR, and Edgerton VR (1998) " Maintenance of myonuclear domain size in rat soleus after overload and growth hormone/IGF-I treatment" J Appl Physiol 84: 1407-1412, 1998
59. McCall GE, Byrnes WC, Dickinson A, Pattany PM, Fleck SJ (1996) " Muscle fiber hypertrophy, hyperplasia, and capillary density in college men after resistance training" J Appl Physiol. 1996 Nov; 81(5): 2004-12.
60. McCormick KM, Thomas DP (1992) " Exercise-induced satellite cell activation in senescent soleus muscle" J Appl Physiol. 1992 Mar; 72(3): 888-93.
61. Moss FP (1968) " The relationship between the dimensions of the fibers and the number of nuclei during normal growth of skeletal muscle in the domestic fowl" Amer. J. Anat. 122: 555.
62. Moss FP, Leblond CP (1970) " Nature of dividing nuclei in skeletal muscle of growing rats" J Cell Biol. 44(2): 459-62.
63. Mozdziak PE, Schultz E, Cassens RG (1997) " Myonuclear accretion is a major determinant of avian skeletal muscle growth" Am J Physiol. 1997 Feb; 272(2Pt1): C565-71.
64. Mozdziak PE, Pulvermacher PM, Schultz E (2000) " Unloading of juvenile muscle results in a reduced muscle size 9 wk after reloading" J Appl Physiol 88: 158-164, 2000
65. Musacchia XJ, Deavers DR, Meininger GA, Davis TP (1980) " A model for hypokinesia: effects on muscle atrophy in the rat" J Appl Physiol. 1980 Mar; 48(3): 479-86.
66. Ogilvie RW, Armstrong RB, Baird KE, Bottoms CL (1988) " Lesions in the rat soleus muscle following eccentrically biased exercise" Am J Anat. 1988 Aug; 182(4): 335-46.
67. Ohira Y, Yoshinaga T, Ohara M, Nonaka I, Yoshioka T, Yamashita-Goto K, Shenkman BS, Kozlovskaya IB, Roy RR, Edgerton VR (1999) " Myonuclear domain and myosin phenotype in human soleus after bed rest with or without loading" J Appl Physiol. 1999 Nov; 87(5): 1776-85
68. Pacy PJ, Evans RH, Halliday D (1987) " Effect of anaerobic and aerobic exercise promoted by computer regulated functional electrical stimulation (FES) on muscle size, strength and histology in paraplegic males" Prosthet Orthot Int. 1987 Aug; 11(2): 75-9.
69. Pavlath GK, Rich K, Webster SG, Blau HM (1989) " Localization of muscle gene products in nuclear domains" Nature. 1989 Feb 9; 337(6207): 570-3.
70. Pullman WE, Yeoh GC (1977) " The role of myonuclei in muscle regeneration: an in vitro study" J Cell Physiol. 1978 Aug; 96(2): 245-51.
71. Ralston E, Hall ZW (1992) " Restricted distribution of mRNA produced from a single nucleus in hybrid myotubes" The Journal of Cell Biology, Vol 119, 1063-1068
72. Rayne J, Crawford GN (1975) " Increase in fibre numbers of the rat pterygoid muscles during postnatal growth" J Anat. 1975 Apr; 119(2): 347-57.
73. Rosenblatt JD, Yong D, Parry DJ (1994) " Satellite cell activity is required for hypertrophy of overloaded adult rat muscle" Muscle Nerve. 1994 Jun; 17(6): 608-13.
74. Roth SM, Martel GF, Ivey FM, Lemmer JT, Tracy BL, Metter EJ, Hurley BF, Rogers MA (2001) " Skeletal muscle satellite cell characteristics in young and older men and women after heavy resistance strength training" J Gerontol A Biol Sci Med Sci. 2001 Jun; 56(6): B240-7.
75. Roy RR, Monke SR, Allen DL, Edgerton VR (1999) " Modulation of myonuclear number in functionally overloaded and exercised rat plantaris fibers" J Appl Physiol. 1999 Aug; 87(2): 634-42.
76. Salleo A, Anastasi G, La Spada G, Falzea G, Denaro MG (1980) " New muscle fiber production during compensatory hypertrophy" Med Sci Sports Exerc. 1980; 12(4): 268-73.
77. Seiden D (1976) " Quantitative analysis of muscle cell changes in compensatory hypertrophy and work-induced hypertrophy" Am J Anat. 1976 Apr; 145(4): 459-65.
78. Sinha-Hikim I, Roth SM, Lee MI, Bhasin S (2003) " Testosterone-induced muscle hypertrophy is associated with an increase in satellite cell number in healthy, young men" Am J Physiol Endocrinol Metab 285: E197-E205, 2003. First published April 1, 2003
79. Sjostrom M, Lexell J, Eriksson A, Taylor CC (1991) " Evidence of fibre hyperplasia in human skeletal muscles from healthy young men? A left-right comparison of the fibre number in whole anterior tibialis muscles" Eur J Appl Physiol Occup Physiol. 1991; 62(5): 301-4.
80. Smith HK, Maxwell L, Martyn JA, Bass JJ (2000) " Nuclear DNA fragmentation and morphological alterations in adult rabbit skeletal muscle after short-term immobilization" Cell Tissue Res. 2000 Nov; 302(2): 235-41.
81. Snow MH (1977) " Myogenic cell formation in regenerating rat skeletal muscle injured by mincing. II. An autoradiographic study" Anat Rec. 1977 Jun; 188(2): 201-17
82. Snow MH (1990) " Satellite cell response in rat soleus muscle undergoing hypertrophy due to surgical ablation of synergists" Anat Rec. 1990 Aug; 227(4): 437-46.
83. Snow MH, Chortkoff BS (1987) " Frequency of bifurcated muscle fibers in hypertrophic rat soleus muscle" Muscle Nerve. 1987 May; 10(4): 312-7.
84. Staron RS, Malicky ES, Leonardi MJ, Falkel JE, Hagerman FC, Dudley GA (1990) " Muscle hypertrophy and fast fiber type conversions in heavy resistance-trained women" Eur J Appl Physiol Occup Physiol. 1990; 60 (1): 71-9
85. Tamaki T, Akatsuka A, Tokunaga M, Ishige K, Uchiyama S, Shiraishi T (1997) " Morphological and biochemical evidence of muscle hyperplasia following weight-lifting exercise in rats" Am J Physiol. 1997 Jul; 273(1 Pt 1): C246-56.
86. Tamaki T, Akatsuka A, Yoshimura S, Roy RR, Edgerton VR (2002) " New fiber formation in the interstitial spaces of rat skeletal muscle during postnatal growth" J Histochem Cytochem. 2002 Aug; 50(8): 1097-111
87. Tamaki T, Uchiyama S, Nakano S (1992) " A weight-lifting exercise model for inducing hypertrophy in the hindlimb muscles of rats" Med Sci Sports Exerc. 1992 Aug; 24(8): 881-6.
88. Timson BF, Bowlin BK, Dudenhoeffer GA, George JB (1985) " Fiber number, area, and composition of mouse soleus muscle following enlargement" J Appl Physiol. 1985 Feb; 58(2): 619-24.
89. Tseng BS, Kasper CE, Edgerton VR (1994) " Cytoplasm-to-myonucleus ratios and succinate dehydrogenase activities in adult rat slow and fast muscle fibers" Cell Tissue Res. 1994 Jan; 275(1): 39-49
90. Vassilopoulos D, Lumb EM, Emery AE (1977) " Karyometric changes in human muscle with age" Eur Neurol. 1977; 16(1-6): 31-4.
91. Wang N, Hikida RS, Staron RS, Simoneau JA (1993) " Muscle fiber types of women after resistance training — quantitative ultrastructure and enzyme activity" Pflugers Arch. 1993 Sep; 424(5-6): 494-502.
92. Winchester PK, Davis ME, Alway SE, Gonyea WJ (1991) " Satellite cell activation in the stretch-enlarged anterior latissimus dorsi muscle of the adult quail" Am J Physiol Cell Physiol 260: C206-C212, 1991
93. Winchester PK, Gonyea WJ (1992) " Regional injury and the terminal differentiation of satellite cells in stretched avian slow tonic muscle" Dev Biol. 1992 Jun; 151(2): 459-72
94. Young RB, Denome RM (1984) " Effect of creatine on contents of myosin heavy chain and myosin-heavy-chain mRNA in steady-state chicken muscle-cell cultures" Biochem J. 1984 Mar 15; 218(3): 871-6.
95. Zilber ML, Litvinova VN, Morozov VI, Pliskin AV, Pshendin AI, Rogozkin VA (1976) " The creatine effect on RNA and protein synthesis in growing culture of chick embryo myoblasts" Biokhimiia. 1975 Jul-Aug; 40(4): 854-60