Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Электрические явления в клеточной энергетике
|
|
Механизм создания АТФ оставался загадкой долгие годы, пока не обнаружилось, что данный процесс по сути своей является электрическим. В обоих случаях: и для дыхательной цепи (набора белков, которые осуществляют окисление субстратов кислородом) и для аналогичного фотосинтетического каскада, — генерируется ток протонов через мембрану, в которую погружены белки. Токи обеспечивают энергией синтез АТФ, а также служат источником энергии для некоторых видов работы. В современной биоэнергетике принято считать АТФ и протонный ток (точнее, протонный потенциал) альтернативными и взаимно конвертируемыми энергетическими валютами. Некоторые функции оплачиваются одной валютой, другие – второй.
К середине XX в. биохимики точно знали, что в бактериях и митохондриях электроны переходят от восстанавливаемых субстратов к кислороду через каскад электронных переносчиков, называемых дыхательной цепочкой. Загадка была в том, каким способом сопряжены перенос электрона и синтез АТФ. На протяжении 10 с лишним лет надежда открыть секрет вспыхивала и вновь угасала. Решающую роль сыграло не преодоление технических трудностей, а концептуальная разработка. Сопряжение оказалось в принципе не химическим, а электрическим. В 1961 г. английский ученый П. Митчелл опубликовал в журнале «Nature» радикальную идею для разрешения биохимической загадки века: хемиосмотическую гипотезу. Идея Митчелла была поистине революционной сменой парадигм, трансформацией концептуальной основы и поначалу вызывала бурные споры.
В 1966 г. Митчелл пишет свою первую книгу «Хемиосмотическое сопряжение в окислительном и фотосинтетическом фосфорилировании». В том же году российские ученые, биофизик Е. Либерман и биохимик В. Скулачев, придумали, как экспериментально подтвердить правоту Митчелла. С помощью синтетических ионов, проникающих через биологическую мембрану, они показали, что дыхание и фосфорилирование, действительно, связаны через протонный потенциал. Еще один серьезный шаг в поддержку Митчелла сделали биофизики биофака МГУ А. Булычев, В. Андрианов, Г. Курелла и Ф. Литвин. Используя микроэлектроды, они зарегистрировали образование трансмембранной разности электрических потенциалов при освещении крупных хлоропластов.
Еще несколько лет споров и дотошных проверок в разных лабораториях по всему свету — и идеи Митчелла, наконец, были признаны. Он был принят в Королевское общество Великобритании (и соответственно, стал сэром), получил множество престижных международных наград, а в 1978 г. был удостоен Нобелевской премии, которая, вопреки традициям, на сей раз была вручена не за открытие нового явления, а за догадку о его существовании.

Хемиосмотическое сопряжение энергии. (а) Общий принцип. (б) Аналогичная электрическая цепь (изображение: www.sciam.ru)
Цепь переноса электрона оказалась не просто связана с мембраной, но вплетена в нее таким образом, что при движении электрона от субстрата к кислороду протоны перемещаются с внутренней поверхности наружу. Мембрана образует замкнутый пузырек, который плохо пропускает протоны, поэтому в результате «выкачивания» протонов генерируется разность потенциалов через мембрану: электрическая отрицательность внутри. Одновременно увеличивается рН: защелачивается среда внутри пузырька. Протоны снаружи оказываются под гораздо более высоким электрохимическим потенциалом, чем внутри, как бы под «давлением» со стороны и электрического потенциала и градиента рН, которые толкают протоны обратно через мембрану внутрь пузырька. Живая клетка использует энергию таких протонов для совершения разных видов работы.
Поразительные успехи рентгеноструктурного анализа белков позволили увидеть полные пространственные структуры отдельных белковых комплексов, входящих в состав дыхательной цепи. Белки цепи переноса электронов, локализованные в мембранах митохондрий, способны менять свой спектр поглощения, получая и отдавая электроны. Микроспектральные методы позволяют проследить последовательность передачи электронов по цепочке белков и выяснить, в каких именно местах часть свободной энергии электронов используется для синтеза АТФ.
Согласно идее Митчелла, для синтеза АТФ из АДФ и фосфата в мембранах митохондрий используется электрическая энергия. Следовательно, если снять разность потенциалов через мембрану, можно предположить, что синтез прекратится. Именно такой эффект был продемонстрирован в ходе экспериментов на искусственных мембранах с использованием специально синтезированных ионов, резко повышающих проводимость мембран для протонов.
Одни из первых экспериментальных доказательств верности гипотезы Митчелла были получены в нашей стране под руководством Е.А. Либермана и В.П. Скулачева. В качестве индикаторов изменений электрического поля на мембране были использованы синтетические ионы, отличающиеся по своей природе и знаку заряда, но сходные в одном: все они легко проникали через фосфолипидную пленку. После многих попыток сложилась следующая изящная экспериментальная модель.
Каплю фосфолипидов, растворенных в органическом растворителе, подносят к небольшому отверстию в тефлоновой пластинке, и оно мгновенно закрывается плоской бимолекулярной пленкой — искусственной мембраной. Тефлоновую пластинку с искусственной мембраной погружают в сосуд с электролитом, разделяя его на два отсека со своим измерительным электродом в каждом. Остается встроить в искусственную мембрану белок, способный генерировать электричество, а в электролит добавить проникающие ионы. Тогда работа белкового генератора, изменяющего разность потенциалов на мембране, приведет к перемещению проникающих ионов через фосфолипидную пленку, что и будет зарегистрировано в виде изменения разности потенциалов между отсеками.
Еще более убедительная экспериментальная модель, позволяющая проводить прямые измерения электрического тока, генерируемого клеточными органеллами и отдельными белками, была разработана и успешно использована Л.А. Драчевым, А.А. Кауленом и В.П. Скулачевым. Частицы, генерирующие электрический ток (митохондрии, хроматофоры бактерий или липидные пузырьки с встроенными в них индивидуальными белками), заставляли слипаться с плоской искусственной мембраной. После этого протонный ток, созданный молекулами-генераторами в ответ на вспышку света или добавление соответствующих химических субстратов, обнаруживался напрямую измерительными электродами по обе стороны искусственной мембраны.
В 1973 г. У. Стокениус и Д. Остерхельт из США открыли необычный светочувствительный белок в мембранах фиолетовых бактерий, обитающих в соленых озерах Калифорнийских пустынь. Этот белок, подобно зрительному пигменту глаза животных – родопсину, — содержал производное витамина А – ретиналь, за что и был назван бактериородопсином. Американские ученые Рэкер и Стокениус изящно продемонтрировали участие бактериородопсина в энергетическом сопряжении. Объединив в модельной фосфолипидной мембране только что открытый светочувствительный белок фиолетовых бактерий с АТФ-синтазой, они получили молекулярный ансамбль, способный синтезировать АТФ при включении света.
В конце 1973 г. академик Ю.А. Овчинников организовал проект «Родопсин» для сравнительного исследования животного и бактериального светочувствительных пигментов. В рамках проекта в лаборатории В.П. Скулачева в МГУ в модельных экспериментах на искусственных мембранах было доказано, что бактериородопсин – белковый генератор электрического тока. Встроенный в искусственную фосфолипидную пленку бактериородопсин направленно транспортировал протоны в ответ на вспышку света. Величина фотопотенциала на мембране превышала 0, 3 В, что заведомо достаточно для энергетического обеспечения синтеза АТФ.
Бактериородопсин оказался на редкость стабильным электрическим генератором: он продолжал работать при нагревании до 100о С и даже в 0, 1 N кислоте. В ходе опытов с бактериородопсином электрическая часть хемиосмотической гипотезы получила свое окончательное подтверждение.
После множества придирчивых проверок теория П. Митчелла была признана абсолютно корректной, и ее рамки были расширены далеко за пределы сопряжения в цепях переноса электрона с синтезом АТФ. Ученому с самого начала было ясно, что циркуляция протонов может поддерживать множество видов работы при посредстве мембранных белков.
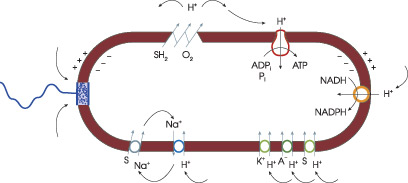
Циркуляция протонов (изображение: www.sciam.ru)
Представим себе, например, белок, транспортирующий субстрат S. Если у белка есть два функциональных места, одно для S, другое для протона, так что поток S сопряжен с потоком протона, до движущая сила для протона оказывается приложенной и к S. Тогда транспорт протона будет не только облегчать перенос S через мембрану, но и действовать как насос, аккумулирующий субстрат внутри пузырька.
У живой клетки есть не только молекулярные генераторы и насосы, но и молекулярные «моторы». Эволюция создала несколько классов белков, способных преобразовывать химическую энергию в механическое усилие. Одни из них используют в качестве топлива гидролиз нуклеотидов, другие – непосредственно ионные градиенты. Есть шаговые белковые моторы, а есть роторные.
Последнее время проблемой преобразования химической энергии в механическую работу активно занялись компьютерные биологи. Они разработали математические модели, описывающие на формальном языке разные типы молекулярных моторов. Принципиальной трудностью, которую им пришлось преодолеть, оказалась невозможность использовать подходы, разработанные ранее для макроскопических моторов, поскольку на работу молекулярных моторов сильное воздействие оказывают термические флуктуации. По этой причине теоретики окрестили белковые моторы «Броуновскими машинами». Тем не менее, в 90-х г.г. XX столетия были разработаны алгоритмы, которые позволили создать ряд имитационных моделей, в частности, мотора бактериального жгутика, механического усилия полимеризующегося волокна, вращающегося мотора АТФ-синтазы.
Главный вывод, к которому пришли исследователи: работа молекулярных моторов вряд ли основана на новых физических или химических принципах, однако, похоже, что для каждого типа белковых моторов придется создавать свое теоретическое описание.
У электрической энергетики живой клетки есть еще одно очень важное свойство. При хемиосмотическом сопряжении энергии и работы не требуется прямого контакта между специфическим белком, создающим разность потенциалов через мембрану, и белком, совершающим какой-то вид работы: поток ионов или метаболитов обеспечит сопряжение двух векторных реакций на расстоянии при условии, что они соответствующим образом ориентированы в одной и той же мембране. Данное свойство успешно используется живыми клетками в митохондриальных сетях, работающих как электрические кабели. Учеными под руководством В.П. Скулачева митохондриальные сети были выявлены у разных типов клеток и исследованы с помощью современных методов электронной микроскопии, прижизненных флуоресцентных зондов, лазерной микрохирургии.
Более того, оказалось, что принцип Митчелла используется группами живых клеток для сопряжения производства энергии одной клеткой с совершением работы в соседней. Необходимое условие такой кооперации – наличие между клетками каналов, пропускающих значительные потоки ионов и метаболитов без утечек во внешнюю среду (см. ст. «Тайны нейроспоры», «В мире науки», №9, 2004 г.).