Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
II. Понятие о раздражимости, возбудимости и возбуждении. Классификация раздражителей
|
|
Раздражимость – это способность клеток, тканей, организма в целом переходить под воздействием факторов внешней или внутренней среды из состояния физиологического покоя в состояние активности. Состояние активности проявляется изменением физиологических параметров клетки, ткани, организма, например, изменением метаболизма.
Возбудимость – это способность живой ткани отвечать на раздражение активной специфической реакцией – возбуждением, т.е. генерацией нервного импульса, сокращением, секрецией. Т.о., возбудимость характеризует специализированные ткани – нервную, мышечные, железистые, которые называются возбудимыми.
Возбуждение– это комплекс процессов реагирования возбудимой ткани на действие раздражителя, проявляющийся изменением мембранного потенциала, метаболизма и т.д. Возбудимые ткани обладают проводимостью. Это способность ткани проводить возбуждение. Наибольшей проводимостью обладают нервы и скелетные мышцы.
Раздражитель – это фактор внешней или внутренней среды действующий на живую ткань.
Процесс воздействия раздражителя на клетку, ткань, организм называется раздражением.
Все раздражители делятся на следующие группы:
1. По природе: а) физические (электричество, свет, звук, механические воздействия и т.д.); б) химические (кислоты, щелочи, гормоны и т.д.); в) физико-химические (осмотическое давление, парциальное давление газов и т.д.); г) биологические (пища для животного, особь другого пола); д) социальные (слово для человека).
2. По месту воздействия: а) внешние (экзогенные); б) внутренние (эндогенные).
3. По силе: а) подпороговые; б) пороговые (раздражители минимальной, силы, при которой возникает возбуждение); в) сверхпороговые (силой выше пороговой).
4. По физиологическому характеру: а) адекватные (физиологичные для данной клетки или рецептора, которые, приспособились к нему в процессе эволюции, например, свет для фоторецепторов глаза); б) неадекватные.
5. Если реакция на раздражитель является рефлекторной, то выделяют также:
а) безусловно-рефлекторные раздражители; б) условно-рефлекторные.
ІІІ. Потенциал покоя (МПП)
Потенциал покоя — относительно стабильная разность электрических потенциалов между наружной и внутренней сторонами клеточной мембраны. Его величина обычно варьирует в пределах 30—90 мВ (в волокнах скелетной мышцы — 60—90 мВ, в нервных клетках — 50—80 мВ, в гладких мышцах — 30— 70 мВ, в сердечной мышце — 80—90 мВ).
ПП играет исключительно важную роль в жизнедеятельности самой клетки и организма в целом, поскольку является основой для возникновения возбуждения (потенциала действия), с помощью которого нервная система воспринимает и перерабатывает информацию, регулирует деятельность внутренних органов и опорно-двигательного аппарата посредством запуска процессов возбуждения и сокращения в мышце. Нарушение процессов возбуждения в кардиомиоцитах ведет к остановке сердца.
Согласно мембранно-ионной теории (Бернштейн, Ходжкин, Хаксли, Катц, 1902— 1952), непосредственной причиной формирования ПП является неодинаковая концентрация анионов и катионов внутри и вне клетки.
Различные ионы распределены неравномерно по обе стороны клеточной мембраны, во-первых, вследствие неодинаковой проницаемости клеточной мембраны для различных ионов, во-вторых — в результате работы ионных насосов, транспортирующих ионы в клетку и из клетки вопреки концентрационному и электрическому градиентам.
Роль проницаемости клеточной мембраны в формировании ПП. Проницаемость клеточной мембраны — это ее способность пропускать воду, незаряженные и заряженные частицы (ионы) согласно законам диффузии и фильтрации. Термин «проводимость» следует использовать только лишь применительно к заряженным частицам. Следовательно, проводимость — это способность заряженных частиц (ионов) проходить через клеточную мембрану согласно электрохимическому градиенту.
Na+ и К+ в покоящейся клетке перемещаются через мембрану согласно законам диффузии, при этом К+ из клетки выходит в значительно большем количестве, чем входит Na+ в клетку, поскольку проницаемость клеточной мембраны для К+ примерно в 25 раз больше проницаемости для Na+.
Органические анионы из-за своих больших размеров не могут выходить из клетки, поэтому внутри клетки в состоянии покоя отрицательных ионов оказывается больше, чем положительных. По этой причине клетка изнутри имеет отрицательный заряд. Интересно, что во всех точках клетки отрицательный заряд практически одинаков. Об этом свидетельствует одинаковая величина ПП при введении микроэлектрода на разную глубину внутрь клетки, как это имело место в опытах Ходжкина, Хаксли и Катца. Гигантский аксон кальмара (его диаметр около 1 мм) в этом опыте находился в морской воде, один электрод вводился в аксон, другой помещали в морскую воду. Заряд внутри клетки является отрицательным как абсолютно (в гиалоплазме клетки содержится больше анионов, нежели катионов), так и относительно наружной поверхности клеточной мембраны. Однако превышение абсолютного числа анионов над числом катионов в клетке чрезвычайно мало. Но этого различия достаточно для создания разности электрических потенциалов внутри и вне клетки.
Главным ионом, обеспечивающим формирование ПП, является ион К+. Об этом свидетельствуют результаты опыта с перфузией внутреннего содержимого гигантского аксона кальмара солевыми растворами. При уменьшении концентрации К+ в перфузате ПП уменьшается, при увеличении концентрации К+ ПП увеличивается. В покоящейся клетке устанавливается динамическое равновесие между числом выходящих из клетки и входящих в клетку ионов К+. Электрический и концентрационный градиенты противодействуют друг другу: согласно концентрационному градиенту К+ стремится выйти из клетки, отрицательный заряд внутри клетки и положительный заряд наружной поверхности клеточной мембраны препятствуют этому. Когда концентрационный и электрический градиенты уравновесятся, число выходящих из клетки ионов К+ сравнивается с числом входящих ионов К+ в клетку. В этом случае на клеточной мембране устанавливается так называемый равновесный калиевый потенциал. Равновесный потенциал для любого иона можно рассчитать по формуле Нернста, а для нескольких по формуле Гольдмана-Ходжкина-Катца
В целом ПП — это производное равновесных потенциалов всех ионов, находящихся внутри и вне клетки и поверхностных зарядов клеточной мембраны.
Вклад Na+ и Сl- в создание ПП. Проницаемость клеточной мембраны в покое для Na+ очень низкая — намного ниже, чем для К+, тем не менее она имеет место, поэтому ионы Na+, согласно концентрационному и электрическому градиентам, стремятся и в небольшом количестве проходят внутрь клетки. Это ведет к уменьшению ПП, так как на внешней поверхности клеточной мембраны суммарное число положительно заряженных ионов уменьшается, хотя и незначительно, а часть отрицательных ионов внутри клетки нейтрализуется входящими в клетку положительно заряженными ионами Na+. Вход Na+ внутрь клетки уменьшает ПП. Что касается Сl -, его влияние на величину ПП противоположно влиянию Na+ и зависит от проницаемости клеточной мембраны для Сl - (она в 2 раза ниже, чем для К+). Дело в том, что Сl -, согласно концентрационному градиенту, стремится и проходит в клетку. Концентрации ионов К+ и Сl - близки между собой. Но Сl - находится в основном вне клетки, а К+ — внутри клетки. Препятствует входу Сl - в клетку электрический градиент, поскольку заряд внутри клетки отрицательный, как и заряд Сl -. Наступает равновесие сил концентрационного градиента, способствующего входу Сl - в клетку, и электрического градиента, препятствующего входу Сl - в клетку. Поэтому внутриклеточная концентрация Сl - равна всего лишь 5—10 ммоль/л, а вне клетки — 120—130 ммоль/л. При поступлении Сl - внутрь клетки число отрицательных зарядов вне клетки несколько уменьшается, а внутри клетки увеличивается: Сl - добавляется к крупным белковой природы анионам, находящимся внутри клетки. Эти анионы из-за своих больших размеров не могут пройти через каналы клеточной мембраны наружу клетки — в интерстиций. Таким образом, Сl-, проникая внутрь клетки, увеличивает ПП. Частично, как и вне клетки, Na+ и Сl - внутри клетки нейтрализуют друг друга. Вследствие этого совместное поступление Na+ и Сl - внутрь клетки не сказывается существенно на величине ПП.
Роль поверхностных зарядов клеточной мембраны и ионов Са2+ в формировании ПП. Наружная и внутренняя поверхности клеточной мембраны несут собственные электрические заряды, преимущественно с отрицательным знаком. Это полярные молекулы клеточной мембраны — гликолипиды, фосфолипиды, гликопротеиды. Фиксированные наружные отрицательные заряды, нейтрализуя положительные заряды внешней поверхности мембраны, уменьшают ПП. Фиксированные внутренние отрицательные заряды клеточной мембраны, напротив, суммируясь с анионами внутри клетки, увеличивают ПП. Роль ионов Са2+ в формировании ПП заключается в том, что они взаимодействуют с наружными отрицательными фиксированными зарядами мембраны клетки и отрицательными карбоксильными группами интерстиция и нейтрализуют их, что ведет к увеличению и стабилизации ПП.
Таким образом, ПП — это алгебраическая сумма не только всех зарядов ионов вне и внутри клетки, но также алгебраическая сумма отрицательных внешних и внутренних поверхностных зарядов самой мембраны.
| , |
Нормальная величина ПП является необходимым условием возникновения процесса возбуждения клетки, т.е. возникновения и распространения потенциала действия, инициирующего специфическую деятельность клетки.
ІІІ. Электротонический и локальный потенциалы (рис.6)
Если воздействовать на клетку раздражителем в размере 1-50% от порогового, клетка ответит электротоничсеким потенциалом – сдвигом МП клетки. Это пассивная реакция клетки на электрический раздражитель; состояние ионных каналов и транспорт ионов не изменяется, либо изменяется совсем незначительно на доли миллисекунд. ЭП не является физиологической реакцией клетки и т.о. не является возбуждением.
Если же на клетку подействовать подпороговым током (50-99% от пороговой величины) – развивается пролонгированный сдвиг МП – локальный ответ. Это активная реакция клетки на раздражитель, однако, состояние ионных и транспорт ионов меняется незначительно. ЛО называют местным возбуждением, т.к. оно не распространяется по мембранам возбудимых клеток, также это не способная к распространению деполяризация мембраны. В основном она обусловлена перемещением ионов Na + в клетку. Вследствие этого уровень поляризации мембраны уменьшается.
Свойства ЛО:
· распространяется с затуханием
· подчиняется закону градуальности (постепенное нарастание или затухание)
· может суммироваться
· нет периода рефрактерности
· имеет фазу деполяризации и реполяризации
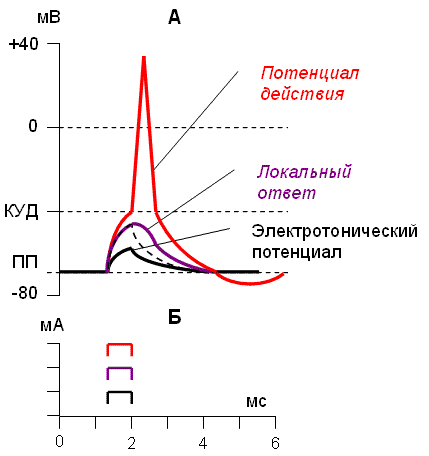 рис. 6
рис. 6