Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Глава 4. · Понятие о липидах и их биологическая роль.
|
|
ОБМЕН ЛИПИДОВ
· Понятие о липидах и их биологическая роль.
· Переваривание и всасывание липидов.
· Метаболизм глицерина.
· Катаболизм жирных кислот.
· Метаболизм кетоновых тел.
· Обмен холестерина.
· Биосинтез липидов.
· Регуляция обмена липидов.
· Нарушения обмена липидов.
· ПОНЯТИЕ О ЛИПИДАХ И ИХ БИОЛОГИЧЕСКАЯ РОЛЬ
Липиды – это относительно разнородная в химическом плане группа органических веществ, нерастворимых в воде и растворимых в некоторых органических растворителях (бензин, бензол, ацетон, эфир и др.). В зависимости от способности к щелочному гидролизу различают омыляемые и неомыляемые липиды. К омыляемым относят жиры (триглицериды, триацилглицерины), воски, фосфолипиды, гликолипиды. По химической природе жиры и воски представляют собой простые эфиры, состоящие из 2-х компонентов – остатка спирта и остатка (остатков) жирной кислоты. Фосфолипиды и гликолипиды являются представителями сложных липидов. В их состав помимо спирта и жирной кислоты входят остаток фосфорной кислоты, аминоспирта или аминокислоты (у фосфатидов) или углеводный компонент (у гликолипидов). К неомыляемым липидам относят стериды. По химической природе это простые эфиры, образованные полициклическими спиртами стеринами и жирными кислотами.
В организме животных содержание липидов колеблется в пределах от 10, 5 до 23 %.
Биологическая роль липидов многообразна. Они являются источниками энергии (при окислении 1 г жира высвобождается 9, 3 ккал энергии), входят в состав клеточных мембран, выполняют защитную функцию, терморегуляторную, для некоторых соединений являются растворителями (для витаминов А, D, E, F, K, Q). При окислении 100 г жира выделяется 107 г эндогенной воды.
· ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЛИПИДОВ
Переваривание различных липидов имеет свои особенности.
Переваривание триглицеридов (жиров).
В ротовой полости корма, содержащие жиры, механически измельчаются, перемешиваются, смачиваются слюной и превращаются в пищевой ком. В составе слюны нет ферментов, расщепляющих триглицериды корма. У взрослых животных жиры проходят через желудок также без особых изменений, поскольку содержащаяся в желудочном соке липаза малоактивна. Величина рН желудочного сока около 1, 5, а оптимальное значение для действия желудочной липазы находится в пределах 5, 5 – 7, 5. Кроме того, липаза может активно гидролизовать только предварительно эмульгированные жиры, а в желудке отсутствуют условия для эмульгирования. Переваривание жиров в полости желудка играет важную роль у новорожденных, рН желудочного сока у них около 5, 0, что способствует перевариванию эмульгированного жира молока желудочной липазой.
Расщепление триглицеридов, входящих в состав корма, у взрослых животных происходит преимущественно в верхних отделах тонкого кишечника, где имеются весьма благоприятные условия для эмульгирования жиров. Наиболее мощным эмульгирующим действием на жиры обладают соли желчных кислот, поступающие в двенадцатиперстную кишку с желчью. Желчные кислоты образуются в печени из холестерина. В желчи содержатся холевая, дезоксихолевая, хенодезоксихолевая, литохолевая кислоты. Желчные кислоты присутствуют в желчи в свободной и конъюгированной (в виде парных соединений соответстветствующей желчной кислоты с глицином или таурином) формах. Соотношение конъюгатов может меняться в зависимости от характера корма: в случае преобладания в них углеводов увеличивается относительное содержание глициновых конъюгатов, а при белковом кормлении – тауриновых. Соли желчных кислот резко уменьшают поверхностное натяжение на границе раздела жир/вода, благодаря чему они не только вызывают эмульгирование, но и стабилизируют уже образовавшуюся эмульсию.
Желчные кислоты выполняют также важную роль в качестве своеобразного активатора панкреатической липазы, под влиянием которой происходит расщепление жиров в кишечнике. Вырабатываемая в поджелудочной железе липаза расщепляет жиры, находящиеся в эмульгированном состоянии. Активирующее влияние желчных кислот на липазу состоит в смещении рН для действия данного фермента с 8, 0 до 6, 0, т.е. до той величины, которая постоянно поддерживается в двенадцатиперстной кишке в ходе переваривания жиров.
Полный гидролиз триглицеридов происходит постадийно: сначала быстро гидролизуюся a- и a½ -связи, а затем медленно идет гидролиз b-моноглицеридов.
В расщеплении жиров может также участвовать и кишечная липаза, однако активность ее невысока. Таким образом основными продуктами, образующимися в тонком кишечнике при расщеплении жиров корма являются глицерин, жирные кислоты, b-моноглицериды.
У взрослых жвачных животных переваривание липидов корма осуществляется главным образом липазами микроорганизмов рубца. Образовавшиеся в ходе расщепления липидов жирные кислоты поступают в кишечник, откуда всасываются и используются для ресинтеза липидов. Кроме того, у жвачных в кишечнике происходит утилизация липидов микроорганизмов за счет панкреатической липазы. Так как микрофлора рубца жвачных восстанавливает экзогенные ненасыщенные жирные кислоты, то снабжение животного ненасыщенными жирными кислотами происходит в основном за счет липидов микроорганизмов.
Переваривание стеридов. Стериды корма эмульгируются под действием тех же факторов, что и триглицериды, после чего расщепляются ферментом холестеринэстеразой до холестерина и высших жирных кислот. Стериды, содержащие остатки насыщенных жирных кислот, расщепляются труднее.
Переваривание фосфатидов. Фосфатиды корма эмульгируются под влиянием тех же веществ, что жиры и стериды. Гидролитическое расщепление фосфатидов происходит под влиянием фосфолипаз А, В, С, D, синтезируемых поджелудочной железой. Каждый фермент действует на определенную сложноэфирную связь фосфатида. Продуктами расщепления фосфатидов являются глицерин, высшие жирные кислоты, фосфорная кислота, аминоспирты (холин, этаноламин) или аминокислота серин.
¯ А1 О
СН2 – О ¾ С // ¾ R1 R1 – остаток насыщенной жирной кислоты
½
½ ¯ А2 О
СН¾ О ¾ С// ¾ R2 R2 – остаток ненасыщенной жирной кислоты
½
½ ¯ C О ¯ D
СН2¾ О ¾ Р// ¾ О ¾ Х Х – азотсодержащий компонент (холин, этаноламин
½ или серин)
ОН
Всасывание липидов. Большинство продуктов расщепления липидов корма всасываются в нижней части двенадцатиперстной и в верхней части тощей кишки. Продукты расщепления липидов всасываются эпителием ворсинок с помощью диффузии, осмоса, активного транспорта и пиноцитоза.
Продукты переваривания липидов корма состоят из мелких частиц жира, ди- и моноглицеридов, высших жирных кислот, глицерина, глицеролфосфатов, азотистых оснований (холина, этаноламина, серина), холестерина, высших спиртов, фосфорной кислоты и др. компонентов. Они распределяются в двух фазах: липидной и мицеллярной (рис. 11). В липидной фазе основными компонентами являются мельчайшие частицы ди- и триглицеридов, в мицеллярной – высшие жирные кислоты, моноглицериды и другие продукты расщепления липидов.
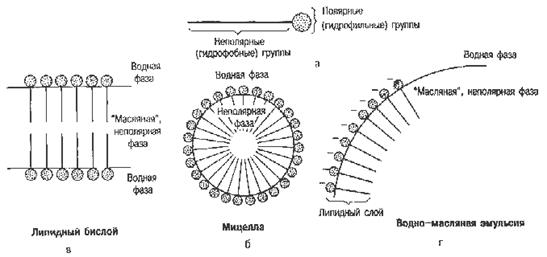
Рис. 11. Строение липидной и мицеллярной фаз.
Продукты переваривания липидов всасываются неодинаково. Легко всасываются глицерин, глицерофосфаты. Фосфорная кислота всасывается в виде натриевых и калиевых солей. Азотистые основания всасываются при участии цитидиндифосфата (ЦДФ). Жирные кислоты с короткой углеродной цепью (менее 10 атомов С) будучи хорошо растворимыми в воде свободно всасываются в кишечнике и поступают в кровь воротной вены, а оттуда в печень без каких-либо превращений в кишечной стенке. В процессе всасывания жирных кислот с длинной углеродной цепью и моноглицеридов, нерастворимых в воде, важная роль принадлежит желчным кислотам. В желчи соли желчных кислот, фосфатиды и холестерин находятся в соотношении 12, 5: 2, 5: 1. Жирные кислоты с длинной цепью и моноглицериды в просвете кишечника образуют с этими соединениями устойчивые в водной среде мицеллы. Структура этих мицелл такова, что их гидрофобное ядро (жирные кислоты, моноглицериды) оказывается окруженным снаружи гидрофильной оболочкой из желчных кислот и фосфатидов. Размеры мицелл примерно в 100 раз меньше самых мелких эмульгированных капель. В составе мицелл высшие жирные кислоты и моноглицериды переносятся с места гидролиза жиров к всасывающей поверхности кишечного эпителия. Здесь происходит распад жировых мицелл, при этом желчные кислоты сразу же поступают в ток крови и через систему воротной вены попадают в печень, откуда они вновь секретируются в составе желчи, т.е. происходит постоянная циркуляция желчных кислот между печенью и кишечником. В желчи содержится небольшая часть желчных кислот (10 – 15 % от общего количества), вновь синтезированных печенью. Основная масса желчных кислот (85 – 90 %) реабсорбируются в кишечнике и повторно секретируются в составе желчи.
В стенке кишечника жирные кислоты и моноглицериды снова превращаются в триглицериды. Здесь также происходит ресинтез фосфатидов (через стадию образования фосфатидной кислоты). Ресинтезированные в эпителиальных клетках кишечника триглицериды и фосфатиды, а также поступивший в эти клетки из полости кишечника холестерин (здесь он может частично этерифицироваться) соединяются с небольшим количеством белка и образуют относительно стабильные комплексные частицы – хиломикроны. Они содержат 2 % белка, 7 % фосфатидов, 8 % холестерина и свыше 80 % триглицеридов. Диаметр хиломикронов колеблется от 100 до 5000 нм. Благодаря большим размерам частиц хиломикроны не способны проникать из эндотелиальных клеток кишечника в кровеносные капилляры и они диффундируют в лимфатическую систему кишечника, а из нее в грудной лимфатический проток, из которого – в кровяное русло. Печень и жировая ткань играют наиболее существенную роль в дальнейшей судьбе хиломикронов. Последние свободно диффундируют из плазмы крови в межклеточные пространства печени (синусоиды). Гидролиз хиломикронов происходит как внутри печеночных клеток, так и на их поверхности. Хиломикроны из-за своих размеров не могут проникать в клетки жировой ткани. В связи с этим триглицериды хиломикронов подвергаются гидролизу на поверхности эндотелия капилляров жировой ткани при участии липопротеидлипазы, которая тесно связана с поверхностью эндотелия капилляров. В результате гидролиза образуются глицерин и жирные кислоты. Часть жирных кислот проникает внутрь жировых клеток, а часть связывается с альбуминами сыворотки крови и уносится с ее током. Покидать жировую ткань с током крови может и глицерин. Схема транспорта липидов указана на рис. 12.
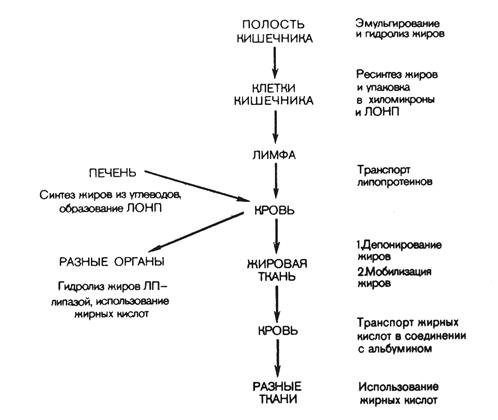
Рис. 12. Транспорт липидов и их использование в тканях.
· МЕТАБОЛИЗМ ГЛИЦЕРИНА
Глицерин образуется в тканях организма животных в результате расщепления триглицеридов под действием тканевых липаз. Функционирование этих ферментов активируется АКТГ гипофиза, адреналином мозгового слоя надпочечников, глюкагоном поджелудочной железы, тирозином щитовидной железы. Посредником в гормональной активации тканевых липаз служит цАМФ. В жировой ткани и печени обнаружены также липазы, отличные от гормоночувствительной липазы.
Глицерин может быть использован в процессах синтеза липидов, глюконеогенеза, а также может окисляться в зависимости от условий до L-лактата или СО2 и Н2О.
В каждом случае использование глицерина идет через глицеролкиназную реакцию, в результате которой образуется глицерол-3-фосфат (Г-3-Ф):
СН2 ¾ ОН СН2 ¾ ОН
| АТФ ½
СН ¾ ОН ¾ ¾ ¾ ¾ ® СН ¾ ОН OН
| глицеролкиназа ½ ½
СН2¾ ОН АДФ СН2¾ О ¾ Р = О
½
ОН
Глицерин Глицерол-3-Фосфат
Далее глицерол-3-фосфат может вовлекаться в липогенез (при взаимодействии с активированными жирными кислотами) или окисляться под действием глицеролфосфатдегидрогеназы с образованием дигидроксиацетонфосфата (ДАФ):
СН2 ¾ ОН СН2¾ ОН
½ НАД+ ½
СН ¾ ОН OН ¾ ¾ ¾ ¾ ¾ ® С = О ОН
½ ½ глицеролфосфатДГ ½ ½
СН2¾ О ¾ Р = О СН2¾ О ¾ Р = О
½ НАДН(Н+) ½
ОН ОН
Глицерол-3 –фосфат ДАФ
ДАФ может использоваться для биосинтеза липидов, в процессе глюконеогенеза или вовлекается в гликолитический путь.
В последних двух случаях ДАФ под действием триозофосфатизомеразы (ТФИ) превращается в глицеральдегид-3-фосфат (ГА-3-Ф):
О
СН2 ¾ ОН С// - Н
½ ½
С = О ОН ¾ ¾ ¾ ¾ ¾ ¾ ¾ ® Н ¾ С ¾ ОН ОН
½ ½ ТФИ ½ ½
СН2¾ О ¾ Р = О СН2 ¾ О ¾ Р = О
½ ½
ОН ОН
ДАФ ГА-3-Ф
В процессе глюконеогенеза ДАФ и ГА-3-Ф реагируют между собой с образованием фруктозо-1, 6-дифосфата (Ф-1, 6-Ф) при участии фермента альдолазы. В дальнейшем Ф-1, 6-Ф под действием фруктозо-1, 6-дифосфатазы превращается во фруктозо-6-фосфат (Ф-6-Ф), который изомеризуется в глюкозо-6-фосфат (Г-6-Ф). Реакция катализируется глюкозофосфатизомеразой (ГФИ). На Г-6-Ф действует глюкозо-6-фосфатаза (фермент присутствует в печени), в результате чего образуется свободная глюкоза.
Н2О Н2О
ДАФ + ГА-3-Ф ® Ф-1, 6-Ф ¾ ¾ ¾ ® Ф-6-Ф ¾ ¾ ¾ ® Г-6-Ф ¾ ¾ ¾ ® глюкоза
Н3РО4 Н3РО4
Для образования одной молекулы глюкозы необходимы 2 молекулы глицерина. Таким образом в организме жиры могут превращаться в углеводы (при недостатке последних в рационе). Однако это превращение носит ограниченный характер.
В случае окислительного пути распада ГА-3-Ф вовлекается в гликолитический путь:
НАД, Н3РО4 АДФ АДФ