Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Окисление b-гидроксибутирата дает 27 молекул АТФ, ацетоацетата – 24 АТФ.
|
|
Кетоновые тела действуют как часть регуляторного механизма с обратной связью, предотвращая мобилизацию жирных кислот из жировых депо, так как последние в высоких концентрациях токсичны для организма.
· ОБМЕН ХОЛЕСТЕРИНА
С химической точки зрения холестерин является полициклическим непредельным одноатомным спиртом, содержащим в своей структуре циклопентанпергидрофенантреновое ядро.

холестерин
Все клетки, содержащие в своем составе ядро, способны синтезировать холестерин. Основным местом синтеза холестерина в организме является печень. Биосинтез холестерина происходит в эндоплазмотическом ретикуломе и цитоплазме клетки. Основным предшественником его является ацетил-КоА.
Первый этап синтеза холестерина начинается так же, как синтез кетоновых тел. Путем конденсации трех молекул ацетил-КоА образуется 3-гидрокси-3-метилглутарил-КоА. В печени митохондриальный запас этого вещества служит предшественником главным образом кетоновых тел, в то время как запас ацетил-КоА в цитоплазме идет на синтез холестерина.
3-гидрокси-3-метилглутарил-КоА под действием редуктазы превращается в мевалоновую кислоту: 3-гидрокси-3-метилглутарил-КоА + 2 НАДФН + 2Н+® мевалоновая кислота + 2 НАДФ+ + КоА-SH
Второй этап начинается с фосфорилирования и декарбоксилирования мевалоновай кислоты, в результате чего он превращается в изопентенилпирофосфат, последний в свою очередь может использьзоватся для построения многих важных веществ организма.
Шесть изопентильных групп затем обьединяются, теряя свои пирофосфатные группы, в результате чего образуется углеводород – сквален.
Третий этап биосинтеза холестерина начинается серией сложных ферментативных реакций, в результате которых линейная молекула сквалена превращается в циклическое соединение – ланостерол, содержащее 4 типичных для стероидов конденсированных кольца.
На четвертой, заключительной стадии ланостерин через серию реакций превращается в холестерин.
Регуляция синтеза холестерина осуществляется на этапе превращения 3-гидрокси-3-метилглутарил-КоА в мевалоновую кислоту. Активность фермента редуктазы, катализирующего эту реакцию, ингибируется конечным продуктом – холестерином. Биосинтез холестерина регулируется также концентрацией специфического белка – переносчика стеролов.
Нарушение регуляции биосинтеза холестерина - это один из факторов, влияющих на развитие атеросклероза.
· Биосинтез липидов
Биосинтез карбоновых кислот. Этот процесс не является обратным пути их окисления, так имеет ряд особенностей:
1. Биосинтез карбоновых кислот протекает в цитоплазме, а их окисление в митохондриях.
2. Ферменты участвующие в синтезе организованы в мультиферментный комплекс, называемой синтетазой жирных кислот, в то время как ферменты, принимающие участи в распаде жирных кислот не склонны к ассоциациям.
3. Промежуточные продукты в биосинтезе жирных кислот связаны с сульфгидрильными группами ацилпереносящего белка (АПБ-SH), а в процессе окисления с сульфгидрильными группами КоА-SH.
4. Источником двухуглеродных фрагментов в процессе синтеза жирных кислот является ацетил-КоА, а непосредственным донором за счет которого осуществляется удлинение углеродной цепи кислоты - малонил-АПБ.
5. Восстановительные реакции в процессе биосинтеза жирных кислот протекают при участии кофермента НАДФН(Н+).
6. Удлинение углеродной цепи (элонгация) в цитоплазме клетки заканчивается на стадии образования пальмитиновой кислоты. Дальнейшее удлинение углеродной цепи осуществляется в митохондриях.
Основным строительным блоком для биосинтеза жирных кислот служит ацетил-КоА, образующийся в митохондриях в результате окислительного декарбоксилирования пирувата, а также в процессе b-окисления жирных кислот. Однако митохондриальная мембрана непроницаема для ацетил-КоА. Поэтому он в митохондриях взаимодействует с оксалоацетатом. В результате реакции, катализируемой цитратсинтазой, образуется цитрат, который переносится в цитоплазму при помощи специальной трикарбоксилат-транспортирующей системы. В цитоплазме цитрат вновь распадается на ацетил-КоА и оксалоацетат. Эта реакция протекает при участии АТФ и КоА-SH.
Перенос ацетил-КоА в цитоплазму может также осуществляться в виде ацетилкарнитина, однако данный путь занимает незначительное место.
В митохондриях ацетил-КоА подвергается карбоксилированию с образованием малонил-КоА. Реакция катализируется ферментом ацетил-КоА-карбоксилазой, содержащей в качестве кофермента карбоксибиотин. Данная реакция лимитирует скорость биосинтеза жирных кислот. Фермент активируется цитратом, а ингибитором является пальмитиновая кислота. Далее ацетил-КоА превращается в ацетил-АПБ, а малонил-КоА – в малонил-АПБ под действием соответственно ацетилацилтрансферазы и малонилацилтрансферазы.
В начале 1-го цикла элонгации происходит конденсация ацетил-АПБ и малонил-АПБ с образованием ацетоацетил-АПБ, содержащим 4 атома С, т.е. на 2 больше, чем в активной уксусной кислоте. Реакция конденсации, катализируемая b-кетоацил-АПБ-синтазой, сопровождается высвобождением СО2. Далее происходит образование b-гидроксибутирил-АПБ при участии b-кетобутирил-АПБ-редуктазы и НАДФН(Н+). Третий этап 1-го цикла элонгации – дегидратация. Здесь образуется транс-кротонил-АПБ. Катализ реакции обеспечивается ферментом еноил-АПБ-дегидратазой. На заключительном этапе 1-го цикла элонгации под действием кротонил-АПБ-редуктазы и НАДФН(Н+) происходит образование бутирил-АПБ.
O O O
// АТФ, СО2 // АПБ-SH //
СН3 – С ~ S – КоА ¾ ¾ ¾ ® НООС – СН2 – С ~ S – КоА ¾ ¾ ¾ ® НООС-СН2– С ~ S – АПБ
Ацетил-КоА АДФ, Н3РО4 малонил-КоА КоА-SH малонил-АПБ
О О
// //
СН3 – С ~ S – КоА + АПБ-SH ¾ ® СН3 – С ~ S – АПБ + КоА-SH
ацетил-АПБ
O O O O
// // // // НАДФН(Н+)
СН3 – С ~ S – АПБ + НООС – СН2 - С ~ S – АПБ ¾ ¾ ¾ ® CH3 – C - CH2 - С ~ S – АПБ ¾ ¾ ¾ ®
CO2, KoA-SH ацетоацетил-АПБ НАДФ
Н О Н О
½ // ½ // НАДФН(Н+)
¾ ¾ ¾ ® CH3 – C - CH2 - С ~ S – АПБ ¾ ¾ ¾ ® CH3 – C = C - С ~ S – АПБ ¾ ¾ ¾ ®
½ Н2О ½ НАДФ
ОН Н
b-гидроксибутирил-АПБ кротонил-АПБ (транс-форма)
O
//
CH3 – CН2 - CH2 - С ~ S – АПБ
бутирил-АПБ
Далее бутирил-АПБ реагирует с малонил-АПБ и реакции повторяются. В конце 2-го цикла элонгации образуется капронил-АПБ, имеющий 6 атомов С. Так повторяется, пока в конце 7-го цикла элонгации не образуется пальмитоил-АПБ, который не может служить субстратом для конденсирующего фермента и поэтому подвергается деацилированию.
О
// Н2О
CH3 – (CH2)14 – C ~ S – АПБ ¾ ¾ ¾ ® CH3 – (CH2)14 – CООН + АПБ-SH
Образование жирных кислот с более длинной углеродной цепью происходит в митохондриях, здесь же образуются и ненасыщенные жирные кислоты из насыщенных. Так, например, из стеариновой кислоты образуется олеиновая, содержащая одну двойную связь в 9 положении.
О O
// О2, НАДН(Н+) или НАДФН(Н+) 10 9 //
CH3 – (CH2)16 – C ~ S – КоА ¾ ¾ ¾ ¾ ¾ ¾ ¾ ¾ ® CH3 – (CH2)7 – CН = СН – (СН2)7 – С ~ S – КоА
Стеароил-КоА оксидаза
2Н2О, НАД или НАДФ олеил-КоА
Линолевая (имеет 2 двойных связи в положениях 9 и 12) и линоленовая (3 двойных связи в 9, 12, и 15 положениях) кислоты не синтезируются в организме и поэтому должны поступать экзогенным путем.
Биосинтез триглицеридов активно протекает в печени, жировой ткани, в молочной железе в период лактации.
Синтез триглицеридов происходит из глицерина и жирных кислот (главным образом стеариновой, пальмитиновой и олеиновой). Путь биосинтеза триглицеридов в тканях происходит через образование a-глицерофосфата (глицерол-3-фосфата) как промежуточного соединения.
Глицерин фосфорилируется за счет АТФ в присутствии фермента глицеролкиназы:
глицерин + АТФ ¾ ¾ ¾ ® глицерол-3-фосфат + АДФ
В жировой ткани и мышцах вследствие низкой активности глицеролкиназы образование глицерол-3-фосфата в основном связано с процессами гликолиза и гликогенолиза. В результате гликолитического распада глюкозы образуется дигидроксиацетонфосфат (ДАФ), который под действием цитоплазматической глицерол-3-фосфатдегидрогеназы способен превращаться в глицерол-3-фосфат (Г-3-Ф):
ДАФ + НАДН(Н+) ¾ ¾ ¾ ¾ ® глицерол-3-фосфат + НАД+
В тех случаях, когда содержание глюкозы в жировой ткани понижено (например, при голодании), образуется лишь незначительное количество глицерол-3-фосфата и освободившиеся в ходе липолиза свободные жирные кислоты не могут быть использованы на ресинтез триглицеридов, поэтому жирные кислоты покидают жировую ткань. Напротив, активация гликолиза в жировой ткани способствует накоплению в ней триглицеридов.
В печени наблюдаются оба пути образования глицерол-3-фосфата.
Образовавшийся тем или иным путем глицерол-3-фосфат последовательно ацилируется 2-мя молекулами ацил-КоА с образованием 1, 2-диацилглицеролфосфата (фосфатидной кислоты), которая гидролизуется фосфатидат-фосфогидролазой до 1, 2-диацилглицерина (1, 2-диглицерида). Это соединение ацилируется третьей молекулой ацил-КоА и превращается в триацилглицерин (триглицерид):
O О O
СН2 ¾ ОН // СН2¾ О ¾ С//¾ R1 //
½ R1 – C ~ S - KoA ½ R2 – C ~ S - KoA
СН ¾ ОН OН ¾ ¾ ¾ ¾ ¾ ¾ ¾ ¾ ¾ ¾ ® СН ¾ ОН OH ¾ ¾ ¾ ¾ ¾ ¾ ¾ ®
½ ½ Г-3-Ф ацилтрансфераза ½ ½ 1- ацилглицерол-3-фосфат-
СН2¾ О ¾ Р = О СН2¾ О ¾ Р = О ацилтрансфераза
½ КоА - SH ½
ОН ОН КоА - SH
Глицерол–3-фосфат 1- ацилглицерол-3-фосфат
О O
// //
СН2 – О ¾ С ¾ R1 СН2 ¾ О ¾ С ¾ R1
½ O ½ O
½ // Н2О ½ // ¾ ¾ ¾ ¾ ®
¾ ¾ ¾ ® СН¾ О ¾ С ¾ R2 ¾ ¾ ¾ ¾ ® CH ¾ O ¾ С ¾ R1
½ фосфатидат- ½
½ O фосфогидролаза ½
½ // CH2 ¾ OH
СН2¾ О ¾ Р ¾ ОH Н3РО4
½
ОН
1, 2 – диацилглицеролфосфат (фосфатидат) 1, 2 – диацилглицерин
О
//
O СН2 – О ¾ С ¾ R1
// ½ O
R2 – C ~ S – KoA ½ //
¾ ¾ ¾ ¾ ¾ ¾ ¾ ¾ ® СН¾ О ¾ С ¾ R2
диацилглицеролацил- ½
трансфераза ½ O
½ //
КоА - SH СН2¾ О ¾ С ¾ R3
триацилглицерин (триглицерид)
В стенке тонкого кишечника синтез триглицеридов возможен из b-моноглицеридов, поступающих из полости кишечника в больших количествах после расщепления экзогенных (пищевых) жиров.
Биосинтез фосфатидов активно протекает в печени, тканях кишечника, почках, мышечной ткани и в меньшей степени в мозге (здесь максимальная скорость синтеза наблюдается в период миелинизации ткани мозга). Высокая скорость синтеза фосфатидов характерна для злокачественных опухолей.
Биосинтез фосфатидилхолина и фосфатидилэтаноламина осуществляется по общему пути. Сначала холин или этаноламин фосфорилируются при участии АТФ с образованием соответственно фосфорилхолина или фосфорилэтаноламина:
СН3 СН3 ОН
½ ½ ½
Н3С ¾ N+ ¾ CH2 ¾ CH2¾ OH + АТФ ¾ ¾ ¾ ¾ ® Н3С ¾ N+ ¾ CH2 ¾ CH2¾ O – Р = О + АДФ
½ холинкиназа ½ ½
СН3 СН3 ОН
холин фосфорилхолин
ОН
½
Н2N ¾ СН2 – СН2 – ОН + АТФ ¾ ¾ ¾ ¾ ® Н2N ¾ СН2 – СН2 – О – Р = О + АДФ
Этаноламин- ½
этаноламин киназа ОН
фосфорилэтаноламин
Образовавшиеся фосфорилхолин и фосфорилэтаноламин взаимодействуют с ЦТФ с образованием ЦДФ-холина и ЦДФ-этаноламина соответственно:
Фосфорилхолин + ЦТФ ¾ ® ЦДФ – холин + Н4Р2О7
Фосфорилэтаноламин + ЦТФ ¾ ® ЦДФ – этаноламин + Н4Р2О7
Далее ЦДФ-холин и ЦДФ-этаноламин реагируют с 1, 2-диацилглицерином, в результате чего образуются соответственно фосфатидилхолин и фосфатидилэтаноламин.
О
//
СН2 – О ¾ С ¾ R1
½ O
½ //
СН¾ О ¾ С ¾ R2 + ЦМФ
ЦДФ – холин + 1, 2 - диглицерид ¾ ¾ ® ½
½ O СН3
½ // ½
СН2¾ О ¾ Р ¾ О ¾ СН2 ¾ СН2 ¾ N+ ¾ СН3
½ ½
ОН СН3
фосфатидилхолин
О
//
СН2 – О ¾ С ¾ R1
½ O
½ //
СН¾ О ¾ С ¾ R2 + ЦМФ
ЦДФ – этаноламин + 1, 2 - диглицерид ¾ ¾ ® ½
½ O
½ //
СН2¾ О ¾ Р ¾ О ¾ СН2 ¾ СН2 ¾ NН2
½
ОН
Фосфатидилэтаноламин
Фосфатидилсерин может образовываться аналогичным образом или в обменной реакции с фосфатидилэтаноламином.
О O
// //
СН2 – О - С - R1 СН2 – О - С - R1
½ O O ½ O
½ // // ½ //
СН¾ О - С - R2 + НО – СН2 – СН - С - OH СН – О – С – R2
½ ½ ¾ ¾ ® ½
½ O NH2 - этаноламин ½ O О
½ // серин ½ // //
СН2¾ О - Р - О - СН2 - СН2 - NН2 CH2 – O – P – O – CH2- CH - C - OH
½ ½ ½
ОН OH NH2
фосфатидилэтаноламин фосфатидилсерин
Таким же образом, а также в обменных реакциях и реакциях взаимопревращения могут синтезироваться и другие фосфолипиды. Фосфатидилэтаноламин может переходить в фосфатидилхолин через промежуточные стадии образования N-метил и N, N-диметилфосфатидилэтаноламинов. В ферментативной системе, необходимой для этого превращения и локализованной в микросомальной фракции печени, в качестве донора метильных групп используется S-аденозилметионин:
Фосфатидилэтаноламин + 3 S-аденозилметионин ® фосфатидилхолин + S-аденозил-
гомоцистеин
· РЕГУЛЯЦИЯ ОБМЕНА липидов
Процессы обмена липидов регулируются нейрогуморальным путем. Центральная нервная система влияет на липидный обмен непосредственно или через железы внутренней секреции. Соответствующие участки коры больших полушарий головного мозга через вегетативные нервные волокна регулируют состав пищеварительных соков, процессы переваривания и всасывания липидов, их биосинтез и мобилизацию. Импульсы, поступающие по симпатическим нервным волокнам, способствуют распаду липидов в жировых депо, а идущие по парасимпатическим – их накоплению.
Центральная нервная система влияет на липидный обмен и через эндокринные железы. Катехоламины (адреналин, норадреналин), глюкагон, тироксин, соматотропный гормон (гормон роста), АКТГ (адренокортикотропный гормон) стимулируют расщепление липидов. Схема регуляции распада липидов указана на рис. 14. Стресс, физическая нагрузка, голодание, охлаждение также усиливают распад липидов через стимуляцию секреции катехоламинов. Инсулин способствует накоплению организмом липидов в результате активации гликолиза в жировой ткани и тормозит освобождение жирных кислот из триацилглицеринов.
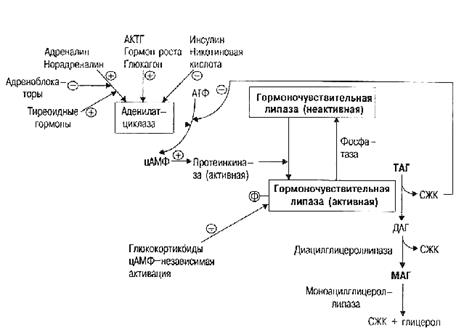
Рис. 14. Схема регуляции липолиза.
· НАРУШЕНИЯ ОБМЕНА липидов
Нарушения обмена липидов может наступать уже в процессе их переваривания и всасывания в желудочно-кишечном тракте. Это может быть вызвано одной из следующих причин:
1. Дефицит панкреатической липазы, связанный с заболеваниями поджелудочной железы.
2. Дефицит желчи в кишечнике, обусловленный заболеваниями печени или желчевыводящих путей.
3. Угнетение ферментных систем ресинтеза триглицеридов в стенке кишечника при его заболеваниях.
Решающая роль в нарушении обмена липидов принадлежит сдвигам в процессах синтеза и распада липидов, которые протекают в тканях, что в конечном счете приводит к нарушению метаболизма.
Важными показателями, характеризующими состояние обмена липидов является содержание в сыворотке крови общих липидов, триглицеридов, фосфолипидов, общего холестерина (табл. 5).
Таблица 5. Содержание липидов в сыворотке крови сельскохозяйственных животных
| Показатели | Единицы измерения | КРС | Лошади | Свиньи |
| Общие липиды | г/л | 2, 5 – 8, 5 | 1, 6 – 2, 6 | 4 – 12 |
| Фосфолипиды | ммоль/л | 0, 91 – 4, 18 | - // - | 1, 17 – 3, 28 |
| Триглицериды | ммоль/л | 0, 03 – 0, 55 | 0, 11 – 1, 4 | 0, 22 – 1, 28 |
| Общий холестерин | г/л | 0, 67 – 2, 88 | 0, 77 – 1, 5 | 0, 9 – 1, 64 |
Содержание общих липидов в сыворотке крови возрастает при желтухах и диабете, триглицеридов – при нефрозах, диабете, гликогенозах, гипотиреозе, беременности, панкреатитах, а снижается при гипертиреозе.
Содержание общего холестерина в сыворотке крови повышено при гепатитах, поражениях почек, гипотиреозе, панкреатитах, гипофункции передней доли гипофиза, ожирении, диабете, беременности, а понижено – при голодании, циррозе, анемии, гиперфункции щитовидной железы.
Обмен липидов нарушается при кетозах. Они возникают при недостатке в рационе углеводов, избыточном кормлении животных концентратами, при сахарном диабете, гепатитах, различных отравлениях. Концентрация кетоновых тел при патологическом кетозе возрастает в крови (кетонемия) и моче (кетонурия) до 20 – 30 ммоль/л (при норме 0, 2 – 0, 6 ммоль/л).
· КОНТРОЛЬНЫЕ ВОПРОСЫ и задания ПО ТЕМЕ:
«ОБМЕН ЛИПИДОВ»
1. Почему запасов липидов в организме хватает на месяц жизни без пищи, а углеводов – только на сутки?
2. Назовите этапы переваривания липидов в желудочно-кишечном тракте. Почему у новорожденных переваривание происходит в желудке, а у взрослых животных – в тонком отделе кишечника?
3. Перечислите факторы, обуславливающие эмульгирование жиров в кишечнике. Какова роль в этом процессе желчных кислот?
4. Укажите основные пути метаболизма глицерина в организме животных. От чего зависит включение глицерина в тот или иной путь метаболизма?
5. Какое значение для организма имеет b-окисление жирных кислот? Почему его так называют?
6. Рассчитайте выход АТФ при окислении предельных жирных кислот, содержащих 16 и 18 атомов углерода.
7. Назовите отличительные особенности процесса биосинтеза жирных кислот по сравнению с их окислением.
8. Какие соединения относятся к кетоновым телам и в чем состоит их биологическая роль? Каковы причины возникновения кетозов?
9. Перечислите гормоны, участвующие в регуляции обмена липидов. В чем заключается их влияние на липидный обмен?
10. Назовите гормоны которые усиливают биосинтез липидов и гормоны, стимулирующие окисление липидов.