Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Морфология и методы исследования вирусов
|
|
Вирус – неклеточная форма жизни, обладающая геномом (РНК или ДНК), но лишенная собственного синтезирующего аппарата и, поэтому, способная к воспроизведению лишь в клетках более высокоорганизованных существ.
По химическому составу и потенциальной патогенности вирусы называют инфекционными нуклеопротеидами. Для вирусов характерны две формы существования: внеклеточная (покоящаяся) и внутриклеточная (репродуцирующаяся, вегетативная). Внеклеточная форма называется вирусной частицей или вирионом. Вирионы состоят из нуклеиновой кислоты, окруженной снаружи белковой оболочкой – капсидом (от лат. сapsa – футляр). Капсид вместе с заключенной в нем нуклеиновой кислотой называют нуклеокапсидом. Морфологическими субъединицами капсида, видимыми в электронный микроскоп, являются капсомеры – белковые субъединицы, состоящие из одной или нескольких молекул белка. Существуют три типа строения капсидов, основанных на расположении морфологических субъединиц (рис. 53):
1. вирионы со спиральной симметрией;
2. вирионы с кубической (икосаэдрической) симметрией;
3. вирионы, имеющие смешанный тип симметрии.
У первого типа капсомеры расположены в виде спирали, нуклеиновая кислота (преимущественно РНК) также скручена в виде пружины, располагаясь между витками белковых молекул. У вирусов с кубической симметрией капсомеры расположены в виде правильного икосаэдра со скрученной в клубок нитью ДНК или РНК. Икосаэдр имеет 20 граней (каждая представляет равносторонний треугольник), 12 вершин. Общее количество капсомеров (N) можно определить по формуле:
N=10 (n-1)2+2, где
n – число капсомеров на одной стороне каждого равностороннего треугольника, которое у различных вирусов варьирует от 2 до 6. Так, аденовирус содержит 252, вирус герпеса – 162 капсомера.
К третьему типу относится вирус осповакцины. Вирус имеет внешнюю оболочку, состоящую из трех слоев, под оболочкой расположены два белковых тела, в центре вириона находится нуклеоид, в состав которого входит ДНК и внутренний белок.
Просто устроенные вирусы, такие как пикорна-, парвовирусы состоят из нуклеокапсида, сложноустроенные вирусы имеют еще дополнительную внешнюю оболочку – суперкапсид или пеплос (производное мембранных структур клетки-хозяина). Форма таких вирионов приближается к сферической. Суперкапсидные белки формируют морфологические субъединицы (пепломеры), которые в электронном микроскопе выглядят в виде шипов (тогавирус, коронавирус, ортомиксовирус и др.). Капсид и суперкапсид защищают вирионы от воздействий окружающей среды, обусловливают избирательное взаимодействие (адсорбцию) с определенными клетками, а также антигенные и иммуногенные свойства вирионов (рис. 53). Размеры вириона колеблются от 20–30 нм (пикорна-, парвовирусы) до 150–250 нм (герпес-, рабдовирусы) и даже 350–400 нм (поксвирусы).
Кроме обычных вирусов, известны и, так называемые, неканонические вирусы: прионы и вироиды. Прионы – это белковые инфекционные частицы, имеющие вид фибрилл размером 10-20х200 нм, они вызывают у животных и человека энцефалопатии в условиях медленной вирусной инфекции (болезнь Крейтцфельда –Якобы, куру и др.). Вироиды – это небольшие молекулы кольцевой, суперспирализованной РНК, не содержащие белка и вызывающие заболевание растений.
Методы исследования вирусов. Для характеристики вирусных частиц широко применяют физические и физико-химические методы. Пользуясь ими, можно определить размер, форму, коэффициент седиментации, коэффициент диффузии, плотность и молекулярный вес как самой вирусной частицы, так и ее компонентов.
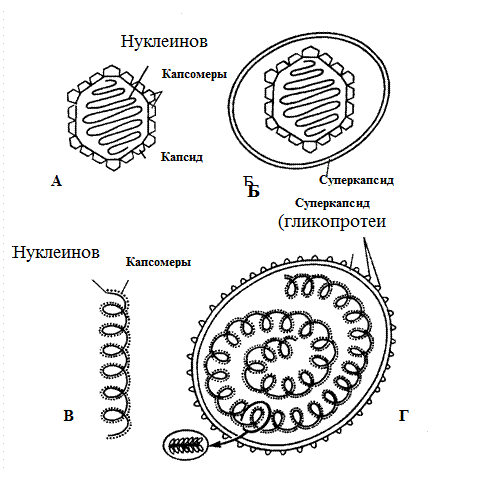 |
Рис. 53. Строение и основные типы симметрии вирусов. А – безоболочечный вирус с икасаэдрическим типом симметрии; Б – оболочечный вирус с икасаэдрическим типом симметрии; В – безоболочечный вирус со спиральным типом симметрии; Г – оболочечный вирус со спиральным типом симметрии. (Медицинская микробиология Под редакцией Покровского В.И. и Поздеева О.К., М., 1998.)
Для определения размеров вирусных частиц используют: фильтрование вируссодержащего материала через мембраны, ультрацентрифугирование, электрофорез, электронную микроскопию.
Фильтрование через коллодиевые мембраны. Метод основан на пропускании вируссодержащего материала через мембраны с известным размером пор. Размер вирусной частицы в данном случае определяется весьма приблизительно.
Осаждение при ультрацентрифугировании. Многие способы определения размеров вирионов основаны на анализе скорости их движения в суспендирующей жидкости. Частицы, взвешенные в жидкости, оседают с разной скоростью, благодаря чему компоненты взвеси можно быстро разделить центрифугированием. Скорость осаждения частицы прямо пропорциональна разности плотности частиц и жидкости, квадрату угловой скорости и квадрату радиуса окружности и обратно пропорциональна вязкости жидкости. Скорость осаждения зависит от формы оседающих частиц. Сферическая частица радиуса r, находящаяся в жидкости с плотностью d0, будет седиментировать в гравитационном поле или поле центробежных сил, если плотность частицы d больше, чем d0.
r=√ 9vŋ 2с(d-d0), где
r – радиус частицы, v – скорость осаждения частицы, ŋ – вязкость среды, с – центробежное ускорение, d – плотность частицы, d0 – плотность жидкости. Это равенство вытекает из формулы Стокса и описывает движение сферических частиц в жидкости при идеальных условиях. Величина S=v/s называется константой седиментации и характеризует поведение данной частицы в данной среде при данной температуре. Константа седиментации выражается в единицах Сведберга, одна единица Сведберга соответствует скорости седиментации в воде при 200С под действием единицы центробежной силы. Так как центробежная сила, плотность среды и ее вязкость могут быть измерены, то можно определить радиус и массу сферической частицы при условии, если мы сможем измерить ее плотность и скорость осаждения в центрифуге. Вирусы хорошо седиментируют в скоростных ультрацентрифугах (60000 об/мин и выше). Используя аналитические роторы, в которых луч видимого или ультрафиолетового света проходит через центрифужные ячейки с прозрачными стенками, можно проводить измерения при движении центрифуги. При седиментации однородной популяции светопоглощающих частиц (вирионов) образуется резкая подвижная граница, положение которой определяют либо непосредственно путем измерения поглощения света, либо по положению области, где показатель преломления жидкости резко изменяется.
Прямое исследование в электронном микроскопе. Электронная микроскопия – наиболее широко применяемый метод определения размеров вирусных частиц. Для этого увеличение на электронных микрофотографиях должно быть откалибровано с использованием внутреннего маркера. С этой целью можно использовать частицы вируса табачной мозаики, имеющие среднюю длину 300 нм и шаг спирали 2, 3 нм; сывороточный альбумин – 5 нм; глобулин – 7 нм; гемоцианин – 23 нм. Метод исключительно быстр, прост и позволяет судить не только о размере вирионов, но отчасти об их форме и характере симметрии.
Методы изучения морфологии вирусных частиц. Детали структуры вируса можно различить только в электронном микроскопе (рис. 54). Широко используется метод негативного контрастирования. Он сводится к смешиванию суспензий вирусных частиц с раствором соли тяжелого металла, нанесению тонкого слоя полученной суспензии на сетку из вольфрамовой пленки и высушиванию полученного препарата. Соль образует плотный слой, на фоне которого материал выглядит сравнительно прозрачным. Обычно соль проникает в различные компоненты вирусной частицы неодинаково, благодаря чему возникает достаточный контраст, способствующий выявлению тончайших деталей структуры вирусной частицы.К числу соединений, наиболее широко применяемых для негативного контрастирования, относятся уксуснокислый и муравьинокислый уранил, кремневольфрамовокислый натрий и молибдат аммония, натриевая или калиевая соль фосфорновольфрамовой кислоты (ФВК).
|
|

А Б
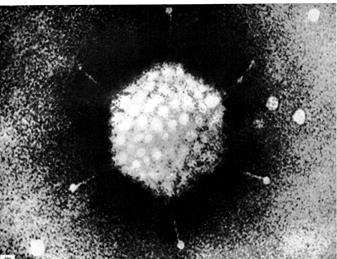
Рис. 54. Вид аденовируса (А) при электронной микроскопии (ув. 600000 раз) и его модель (Б). (Авакян А.А., Быковский А.Ф., М., Атлас анатомии и онтогенеза вирусов человека и животных, 1970).
Важные сведения о структуре и морфогенезе вирусов дает также метод тонких срезов, используемый для изучения препаратов вирусов, находящихся в осадке. Для того чтобы установить локализацию специфических белков в вирусной частице используют вспомогательные методы, например радиоавтографию, обработку тонких срезов мечеными антителами.
В ряде случаев используется действие на вирионы поверхностно-активных веществ. При этом оценивают формы, оставшиеся после обработки и делаются выводы о том, какие компоненты из структуры были удалены.
Для изучения вирусных нуклеиновых кислот используют методы гибридизации нуклеиновых кислот.
Изучение физико-химических свойств вирусов
Электрофорез. Электрофоретические методы позволяют определить специфическую физическую характеристику вирусной частицы – относительную электрофоретическую подвижность под влиянием электрического поля. В противоположность коэффициентам седиментации и диффузии эта величина практически не зависит от массы частицы и основывается главным образом на суммарном заряде поверхности частицы. Расчет константы электрофоретической подвижности производится по формуле:
U= χ.q.s,
i.t
где χ – удельная проводимость, в Ω -1.см-1; q – поперечное сечение электрофоретической колонки, в см; s – путь, пройденный частицей; i – сила тока в А; t – время пробега в сек. Подвижность выражается в см2/вольт/сек.
Классический метод электрофореза, или метод движущейся границы Тизелиуса, широко используемый для анализа белков применяется для физико-химической характеристики вирусных частиц – определения изоэлектрической точки и электрофоретической подвижности.
Фронтальный электрофорез – единственный метод, позволяющий определить электрофоретическую подвижность и изоэлектрическую точку с очень высокой точностью. Кроме того, этот метод широко используется при исследовании гомогенности и степени чистоты вирусных препаратов.
Еще более разрешающий метод – метод зонального электрофореза в градиенте плотности и в гелях. Его используют не только для изучения специфической электрофоретической подвижности вирусов, их дифференцировки и идентификации, но и для выделения генетически однородных штаммов и изучении изменчивости микроорганизмов.
Методы фракционирования вирусов. Эти методы позволяют проводить дезинтегрирование вирусных частиц на отдельные компоненты и их фракционирование. Выделенные компоненты в дальнейшем можно подвергнуть биохимическому анализу с целью изучения их тонкой структуры и свойств. Фракционирование компонентов вирусной частицы осуществляется при помощи центрифугирования в зональном и равновесном градиентах плотности, зонального электрофореза и хроматографии.
Хроматография. Для очистки и фракционирования вирусов применяют три типа хроматографии: адсорбционную, ионообменную и молекулярно-ситовую.
Метод адсорбционной хроматографии основан на различной степени адсорбции компонентов смеси при фильтровании через неподвижный твердый адсорбент. Решающее значение имеют поверхностные свойства вирусной частицы и адсорбента, а также состав буфера, в котором суспендирован вирус. При элюировании соответствующим буфером вирусные частицы можно отделить от примесей. В качестве адсорбентов для наполнения хроматографических колонок в вирусологической практике чаще всего используют фосфат калия и гидроксилапатит.
При ионообменной хроматографии вирусы пропускают через ионообменник. Ионообменниками называют такие соединения, которые содержат фиксированные функциональные группы и подвижные противоионы. Последние могут обратимо обмениваться с другими ионами того же заряда, не изменяя физические свойства нерастворимой матрицы. Ионообменниками могут быть органические и неорганические соединения. В качестве матрицы могут быть использованы алюмосиликаты, синтетические смолы, полисахариды, белки и целлюлоза.
Метод молекулярно-ситовой хроматографии основан на способности пористых материалов, работающих по принципу «обратных молекулярных сит», разделять смесь веществ по размеру и молекулярному весу компонентов. В качестве таких пористых материалов применяют гранулированные гели полисахаридов (сефадексы, агароза), полиакриламида (биогели) и пористое порошковое стекло. Метод подобного фракционирования обычно называют молекулярно-ситовой хроматографией, молекулярной фильтрацией или гельфильтрацией.