Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Фагоцитоз, опсониндер
|
|
Фагоцитоз – корпускулалардың, бактериялардың немесе ү лкен макромолекулярлы кешендердің фагоцитпен жұ тылуы. Фагоцит жасушалары: нейтрофилдер жә не моноциттер/ макрофагтар. Эозинофилдер де фагоцитозғ а қ атыса алады (антигельменттік иммунитет кезінде нә тижесі кө бірек болады.). Фагоцитоз ү рдісін фагоцитоз нысанын бү ркейтін опсониндер кү шейтеді.
Қ андағ ы лейкоциттердің 5− 10%-ын моноциттер, ал 60− 70 %-ын нейтрофилдер қ ұ райды. Моноциттер тінге тү се отырып, тіндік макрофагтар популяциясын қ алыптастырады: купфер жасушалары (немесе бауырдың жұ лдызшалы ретикулоэндотелиоциттері), сү йек тінінің остеокласттары, альвеолярлы жә не интерстицийлік макрофагтар, ОЖЖ микроглиясы.
Фагоцитоз ү дерісі. Фагоциттер хемоаттрактанттардың (микробтардың заттектері, комплементтің белсендірілген компоненттері (C5a, C3a) жә не цитокиндер) ә серіненфагоцитоз нысанына бағ ыт алып жылжиды. Фагоциттің плазмалеммасы бактерияларды немесе басқ а корпускулаларды жә не ө зінің бұ зылғ ан жасушаларын ұ стайды. Содан кейін фагоциттің нысаны плазмалеммамен қ оршалады да, мембраналық везикула (фагосома) фагоциттің цитоплазмасына енеді. Фагасома мембранасы лизасомамен бірігеді де фагоциттелген микроб бұ зылады, рН− 4, 5-ке дейін қ ышқ ылданады; лизасома ферменттері белсене тү седі. Фагоциттелген микроб лизосома ферменттерінің, катионды нә руыздар дефенсинінің, G катепсинінің, лизоцимнің жә не басқ а да факторлардың ә серінен ыдырайды. Оттегілік жарылыс кезінде фагоцитте оттегінің улы антимикробты тү рлері – Н2О2 сутек асқ ынтотығ ы, О2 супероксиданион, ОН гидроксилді радикал, синглетті оттегі пайда болады. Сонымен қ атар азот тотығ ы жә не NO радикалының да антимикробты ә сері бар.
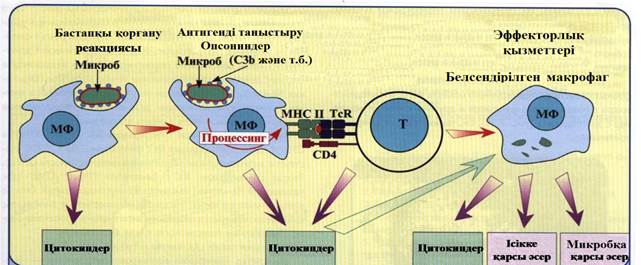
7-сурет. Макрофактардың иммунитеттегі орталық рө лі
Макрофагтар иммунокомпетентті жасушалармен байланыспай тұ рып қ орғ аныш функциясын 7-сурет) атқ арады (бейспецификалық резистенттілік). Макрофагтордың белсенуі фагоциттелген микробтардың бұ зылуынан, оның процессингінен (қ айта ө ң делу), Т-лимфоциттерге презентациясынан (таныстырылуы) кейін болады Иммундық жауаптың соң ғ ы сатысында Т-лимфоциттер макрофагты белсендіретін цитокиндер бө леді (жү ре пайда болғ ан иммунитет). Белсендірілген макрофаг антидене жә не белсенді комплемент жү йесімен қ осылып, фагоциттелген микробтарды бұ за отырып, жоғ ары нә тижелі фагоцитозды жү зеге асырады (иммундық фагоцитоз).
Фагоцитоз ұ стап алынғ ан микробтардың жойылуы нә тижесінде − аяқ талғ ан жә не микробтар тіршілігін жоймағ ан жағ дайда − аяқ талмағ ан болуы мү мкін. Аяқ талмағ ан фагоцитоздың мысалына гонококктардың, туберкулез таяқ шаларының жә не лейшманиялардың фагоцитозы жатады.
|
Грамоң бактериялар (стафилококктар, стрептококктар, пропионбактериялар жә не т.б.) капсула тә різді жамылғ ы тү ріндегігуморалдық факторларды ө зіне тә н еместү рдеадсорбциялауғ а қ абілетті. Осының ә серінен олар грамтеріс бактерияларғ ақ арағ анда қ арқ ынды фагоциттеледі (сондық тан кө птеген авторлар фагоцитозды бағ алау кезінде грамоң бактерияларды қ олдануды дұ рыс деп есептемейді).Осылайша,
|
шартты тү рде иммуноглобулиндік жамылғ ы деп аталатын қ ан сарысуынан ө ң делген стафилококктар капсула тә різдес жамылғ ы тү рінде (8, 9-сурет) сарысу нә руыздарын (иммуноглобулиндер, комплемент, альбумин жә не т.б.) ө зіне тә н емес тү рде адсорбциялайды (А.С.Быков, 1978). Опсониндер иммунды жабысу нә тижесінде, фагоциттелетін объектінің гидрофобты қ асиеттерінің кү шеюінен фагоциттердің жоғ арғ ы беткейімен жә не басқ а жасушалармен ә рекеттесе алады, мысалы эритроциттермен (10, 11-сурет).
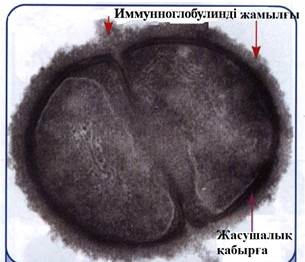
8-сурет. Стафилококктардың иммуноглобулиндік
жамылғ ысының ультражұ қ а электрограмма
кескіні.

9-сурет. Қ алыпты адамның иммуноглобулиндік препаратымен ө ң делген
стафилококктың жә не ішек таяқ шасының ультражұ қ а кескін электрограммасы.
IgG жиналуына байланысты тек стафилококкта ғ ана
иммуноглобулиндікжамылғ ы пайда болады.
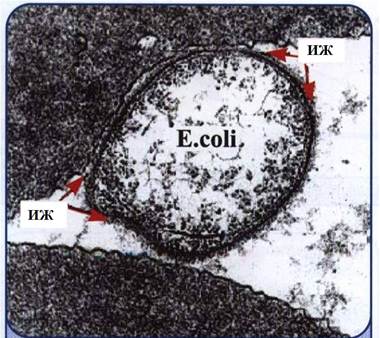
10-сурет. E.coli эритроциттеріне иммуноглобулиндік
жамылғ ыарқ ылы иммундық жабысу.

11-сурет. Қ ан сарысуымен опсонинделген
S.aureusфагоцитозы. Ультражұ қ акесіндінің электрограммасы.
 |
ТЖ жасушалары (normal killers. natural killers − табиғ и жендет-жасушалар) − цитокиндермен бірлесе отырып, вирус жұ қ қ ан жасушаларғ а, қ артайғ ан немесе ісік жасушаларына қ арсы ө зіне тә н емес цитоуыттылық кө рсетеді.
ТЖ жасушалары лектиндік тану немесе антиденеге-тә уелді цитоуыттылық негізінде нысана-жасушаларды жойып отырады. ТЖ жасушалары МНС-І класын экпрессияламайтын (кейбір жағ дайда) нысана− жасушаларын жойып отырады. ТЖ жасушалардың цитоуыттылық ә рекеті перфоринге тә уелді механизмдермен себептес жә не цитотуытты лимфоциттердің ә рекетіне ұ қ сас. ТЖ жасушаларының бө где антигендері бар жасушаларғ а бекінген антиденелердің Fс-фрагментімен қ осылуы кезінде жасушалық антиденегетә уелді цитоуыттылық ә сері дамып, нысана-жасушалардың ө ліміне ә келеді.
ТЖ жасушаларының екі негізгі субпопуляциялары бар: CD56+ /CD16‾ жә не CD56‾ /CD16+. CD56+ /CD16‾ фенотипті ТЖ субпопуляциясы антиденегетә уелді жасушалық цитоуыттылық қ а қ атысады. CD56‾ /CD16+ фенотипті ТЖ екінші субпопуляциясы (Pit-жасушлары) бауыр синусоидында орналасқ ан. Дә л осындай жасушалар жатырда да бар. Тағ амдық антигендермен жә не ұ рық антигендерімен белсендірілетін бұ л жасушалар барлық лимфоциттерді жоя алады жә не осы антигендерге тө зімділікті қ алыптастырады.
Сонымен қ атар LAK-жасушалары мен γ δ Т-жасушалары болып ажыратылады. LAK-жасушалары (лимфокин-белсендірілген жендет) ТЖ жасушаларының аздағ ан субпопуляциясынан туындайды. Ол ісік жасушаларына цитоуыттық ә сер береді, ә сіресе интерлейкиндердің (ИЛ2, ИЛ1, ИЛ6, ИЛ15), интерферондардың, IНФα жә не т.б. цитокиндердің ә серінен. LAK− жасушалары Т-лимфоциттерді жә не эозинофилддерді белсендіреді. LAK-терапиясын меланома жә не бү йрек карциномасын емдеуде қ олданады. γ δ Т-жасушалары немесе ТЖ жасушалары
Т-жасушалардың аздағ ан субпопуляциясы болып табылады: ТЖ-жасушасының мү мкін болатын аналогы. Ол барьерлі ағ заларда жә не сілемейлі қ абық тарда орналасқ ан. Туберкулез қ оздырғ ыштары мен оппортунистік жұ қ паларды элиминациялайалады.


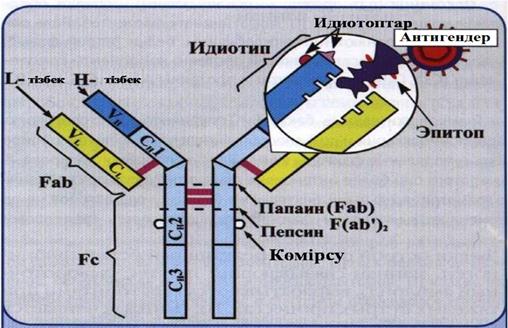
12-сурет. Ig G1 молекуласының қ ұ рылысы.
Иммуноглобулиндердің мономері дисульфидтік арақ атынаспен байланысқ ан екі ауыр (Н-тізбектен) жә не екі жең іл (L-тізбектен) тұ рады (12-сурет). Бұ л тізбектер константты жә не вариабельді аумақ тардан тұ рады. Ауыр тізбектің тү рі бойынша иммуноглобулиндердің 5 класы ажыратылады: Ig G, Ig M, Ig A, Ig D, Ig E. Папаин иммуноглобулин молекуласын екі антигенбайланыстырушы фрагменттерге бө леді − Fab (fragment antigen binding) жә не Fc (fragment cristalizable). Иммуноглобулиннің Fab фрагментінің антиген байланыстырушы аумағ ы (антиденелердің белсенді аумағ ы) Н-жә не L-тізбектердің гипервариабельді аумағ ынан қ ұ ралғ ан; ол антигеннің эпитоптарын байланыстырады. Белсенді орталық та белгілі антиген эпитоптарына арнайы комплементарлы аймақ тар болады. Ғ сфрагменті комплементті байланыстырады (антиген-антидене кешені пайда болғ анда), жасуша мембранасымен ә рекеттеседі жә не IgG-дің плацента арқ ылы тасымалдануына қ атысады.
Дисульфидтікарақ атынаспен байланысқ ан антидененің ық шам қ ұ рылымы − домендер деп аталады. Сонымен, IgG-дің: Fab-фрагментінің N-ұ штық аймағ ында орналасқ ан жең іл (VL) жә не ауыр (VH) тізбектердің вариабельді V-домені; жең іл (CL) тізбектердің константты аумақ тарының С-домені; ауыр (CH1, CH2, CH3) тізбектердің константты аймақ тарының С-домені болып ажыратылады. СН2-доменінде комплемент байланыстырушы аумақ болады.
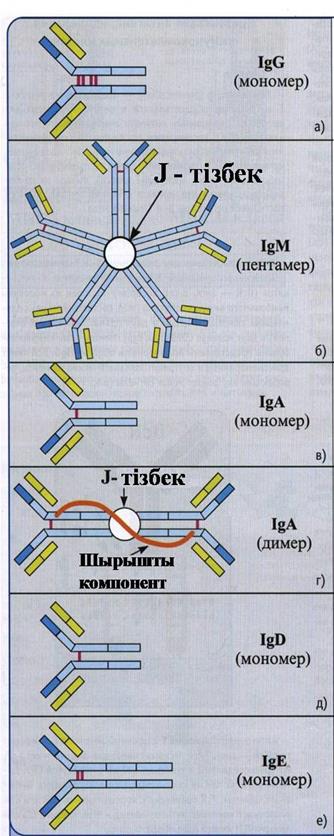
a) IgG (G иммуноглобулині антидене қ ан сарысуының 80% қ ұ райды; 4 топша IgG 1, IgG 2, IgG 3, IgG 4)
1.Топшасына қ арай жартылай ыдыратудың 7-23 кү ндік кезең і.
2.Мономер; аймақ ты байланыстырушы -2 эпитоп
3.Fc-фрагменті комплименттің белсендірілуінің классикалық жолына қ атысуы мү мкін.
4.Fc-фрагменті макрофактармен, нейтрофилдер мен NK-жасушаларымен байланысуы мү мкін
IgM (М иммуноглобулині антидене қ ан сарысуының 10% қ ұ райды)
1.Жартылай ыдырау кезең і-5 кү н аралық.
2.Иммундық жауап кезінде ө ндірілетін бірінші антидене.
3.Пентамер: 10эпитоп байланыстырушы аймақ тар.
4.Fc-фрагментті комплименттің белсендірілуінің классикалық жолына қ атысуы мү мкін.
5.Мономерлі формалар В-лимфоциттің жоғ арғ ы бетінде IgM тү рінде болады.
IgА (А иммуноглобулині антидене қ ан сарысуының 9 % қ ұ райды; 2топша – IgА1, IgА2) жартылай ыдырау кезең і -5 кү н шамасында
Секреторлық IgА (s IgА) сілемейлі қ абық та, сілекейде, кө з жасында, уызда, ана сү тінде кездесіп, вирустар мен бактерияларды бө гейді. Димер сілемейлі компанеттен тұ рады, антиденені ферменттердің ыдырауынан қ орғ айтын: 4 эпитоп байланыстырушы аймағ ы бар.
IgD (иммуноглобулин қ ан D сарысуының 0, 2 % антиденесін қ ұ райды).
1.Мономер; 2 эпитоп байланыстырушы аумағ ы бар.
2. В-лимфоциттің жоғ арғ ы бетінде mIg тү рінде кездесіп, активациясы мен супрессиясын байланыстырады. (мономерлі IgM -қ атар).
IgE (иммуноглобулин Е қ ан сарысуының 0, 002 % антиденесін қ ұ райды)
1.жартылай ыдырау уақ ыты 2 кү н шамасында.
2.мономер; 2 эпитоп байланыстырушы аумағ ы бар.
3.Паразитке қ арсы иммунитетке қ атысады жә не аллергендерге жауап береді. Антидененің Fс-фрагментімес жасушасымен жә не базафилдермен ибайланысады, кейіннен аллергендермен ә рекеттескенде аллергиялық реакция тудырады.
13-сурет. а − д. G, M, A, D, E класты иммуноглобулиндердің қ ұ рылысының сызбасы мен қ ызметтері.