Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Развитие тычинки. Микроспорогенез и формирование мужского гаметофита. Вскрывание пыльника
|
|
Тычинки закладываются на цветоложе в виде бугорков после появления на нем зачатков будущего околоцветника. Из бугорка, первоначально состоящего из однородных тонкостенных клеток, формируется пыльник, а тычиночная нить развивается путем интеркалярного роста перед распусканием цветка.
Анатомическая дифференциация пыльника начинается, когда бугорок приобретает четырехлопастные очертания поперечного сечения (рис. 236 А, В). Под эпидермой в каждой лопасти обособляются крупные инициальные клетки. Их тангентальные (периклинальные) деления приводят к образованию 2 слоев: наружного — париетального — и внутреннего, дающего начало археспорию, или спорогенной ткани (рис. 236 Г, Д). Клетки археспория либо непосредственно становятся микроспороцитами, либо предварительно делятся митотически. Каждая лопасть формирующегося пыльника с археспорием соответствует одногнездному микроспорангию. Эпидерма и производные париетального слоя составляют стенку пыльника. Эпидерма имеет типичное для этой ткани строение. Наружные стенки ее клеток обычно утолщены и покрыты кутикулой, а иногда и восковым налетом. Клетки содержат хромопласты.
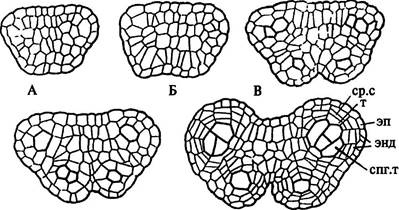
| Рис. 236. Начальные стадии развития пыльника нивяника. Обозначения: спг.т — спорогенная ткань, ср.с — средний слой, т — тапетум, энд — эндотеций, эп — эпидерма |
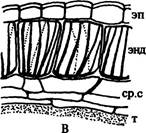
Париетальный слой вследствие периклинальных делений его клеток образует эндотеций, расположенный под эпидермой, и несколько слоев паренхимных клеток. Эндотеций называют фиброзным слоем, если оболочки его клеток с внутренней стороны имеют разнообразные утолщения, способствующие вскрыванию пыльника (рис. 237 В). Если он не участвует в этом процессе, то оболочки его клеток не утолщены. Самый внутренний слой расположенных под эндотецием паренхимных клеток, выстилающий каждое гнездо пыльника, составляет тапетум (у некоторых растений он может состоять из 2 и более слоев) (рис. 237 В). Тапетум может развиваться не только из производных париетального слоя, но и из наружного слоя археспория.
Молодые клетки тапетума одноядерные, позднее число ядер в них увеличивается вследствие митотических делений, нередко они соединяются между собой, образуя крупные полиплоидные ядра. У покрытосеменных встречается 2 типа тапетума. Первый тип - секреторный тапетум - состоит из железистых клеток, экскретирующих вещества в гнезда пыльника. Второй тип - периплазмодий - возникает вследствие растворения оболочек клеток и слияния их содержимого в общую массу, проникающую в гнездо пыльника. Содержащиеся в тапетуме питательные вещества впоследствии используются на развитие пыльцевых зерен. Находящиеся в гнездах пыльника диплоидные микроспороциты (материнские клетки микроспор) первоначально плотно соединены. Затем они расходятся и покрываются каллозой, изолирующей их от остального содержимого пыльцевого гнезда. Делясь редукционно, каждый микроспороцит образует тетраду микроспор.
| 
| 
|
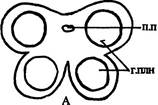
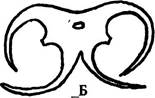
| Рис. 237. Строение пыльника: А — невскрывшийся пыльник; Б — вскрывшийся пыльник; В - стенка пыльника. Обозначения: г.плн — гнезда пыльника, п.п — проводящий пучок, ср.с — средние слои клеток, т — тапетум, энд — эндотеций, эп — эпидерма |
Стенки, отделяющие одну микроспору от другой, могут развиваться двояко. При сукцессивном, или последовательном, типе первое, редукционное, деление мейоза заканчивается цитокинезом, в результате образуется диада клеток, а после второго, митотического деления - тетрада. При одновременном, или симультанном, типе цитокинез происходит после второго деления, и сразу образуется тетрада микроспор. Сукцессивный тип свойствен большинству однодольных, а симультанный распространен у двудольных растений.
Чаще всего у покрытосеменных встречаются тетраэдрические и билатеральные тетрады (рис. 204).
Разъединение собранных в тетрады микроспор происходит после растворения каллозы и собственной оболочки микроспороцита. К этому времени каждая микроспора имеет 2 оболочки: наружную - экзину и внутреннюю - интину.
Микроспора представляет собой одноядерную гаплоидную клетку, из которой внутри пыльника развивается мужской заросток - пыльцевое зерно, или пылинка. Зрелое пыльцевое зерно, покидающее пыльник, может быть двухклеточным или трехклеточным. В первом случае оно состоит из крупной вегетативной, или сифоногенной, клетки и погруженной в нее маленькой генеративной, или спермиогенной, клетки, которая делится на 2 спермия только в пыльцевой трубке. Во втором случае спермин образуются уже в самом пыльцевом зерне (рис. 238 Г, Д). Спермии представляют собой клетки, имеющие сравнительно крупное ядро, плотную цитоплазму и очень тонкую прозрачную оболочку. У разных растений они сильно варьируют по размерам и форме. Спермии могут быть более или менее шаровидными, эллиптическими, серповидными, веретеновидными, червеобразными, спирально закрученными и др.
Как и микроспора, пыльцевое зерно имеет 2 оболочки: экзину и интину. Пыльцевые зерна некоторых растений имеют еще третью оболочку - перину, расположенную поверх экзины. Она возникает за счет тапетального периплазмодия. Перина соединена с экзиной неплотно и легко разрушается.
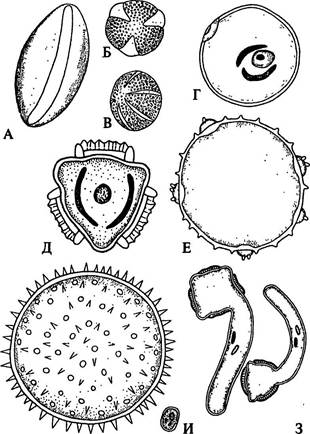
| Рис. 238. Типы пыльцевых зерен: А, Б, В — одно-, трех- и многобороздные пыльцевые зерна магнолии, пиона, гименократера; Г — однопоровое пыльцевое зерно многолетней пшеницы; Д — трехпоровое зерно кок-сагыза; Е — многопоровое зерно тыквы; Ж — многопоровое зерно мальвы; 3 — трех-и четырехпоровые зерна диплоидной и триплоидной форм табака; И — пыльцевое зерно незабудки |
Экзина имеет несколько слоев, самый наружный из них часто скульптурирован и снабжен всевозможными выростами в виде гребней, бугорков, шипиков, складок и т. п. (рис. 238).
Экзина, состоящая из споропоменина, очень устойчива к разным воздействиям, в том числе химическим. Она имеет утонченные места - апертуры, иногда достигающие интины. По характеру апертур, через которые прорастают пыльцевые трубки, различают бороздные (рис. 238 А-В) и поровые (рис. 238 Е, Ж) пыльцевые зерна, число борозд и пор варьирует у разных растений от одной до многочисленных.
Интина, содержащая целлюлозу и пектиновые вещества, обладает эластичностью. Это имеет важное значение, так как при прорастании пыльцевого зерна она, сильно растягиваясь, образует пыльцевую трубку, развивающуюся из сифоногенной клетки. Через апертуру пыльцевая трубка выходит наружу (рис. 238 3).
Пыльцевые зерна покрытосеменных несомненно гомологичны микрогаметофитам других разноспоровых высших растений, но более редуцированы. Их сходство с пыльцевыми зернами голосеменных проявляется в едином плане строения оболочек и образовании только 2 спермиев. Однако у покрытосеменных микрогаметофиты редуцированы в большей степени, так как при их развитии от стадии микроспоры до образования спермиев происходит только 2 митотических деления вместо 5, свойственных, например, большинству хвойных. Проталлиальные клетки, характерные для голосеменных, у покрытосеменных полностью утрачены.
Морфологически пыльцевые зерна весьма разнообразны (рис. 238). Очень мелкие (менее 10 мкм в поперечнике), практически неразличимые простым глазом пыльцевые зерна - у незабудки, некоторых видов пеперомии, очень крупные (свыше 200 мкм) - у мальвы, алтея, ипомеи.
Зрелые пыльцевые зерна не всегда расположены поодиночке. Нередко они остаются собранными в тетрады, как у грушанки, вереска, брусники; диады, как у рябчика, и полиады, или массулы, как у некоторых представителей семейства ароидных. У многих орхидей все находящиеся в пыльцевом гнезде пыльцевые зерна склеены висцином в крупное компактное образование - поллиний, который вместе с ножкой и прилипальцем, прилипающим к посетившему цветок насекомому, называют поллинарием.
Морфология пыльцевых зерен, строение их оболочек представляют собой очень устойчивые видоспецифические признаки, широко используемые в систематике.
Освобождение пыльцы происходит при вскрывании пыльника. Пыльники, в которых гнезда попарно сближены, вскрываются обычно двумя продольными щелями, проходящими вдоль борозд, разделяющих гнезда, так что оба гнезда вскрываются одновременно (рис. 237 Б). При продольном вскрывании щель обычно появляется с внутренней стороны пыльника, и пыльца высыпается внутрь цветка. Такой пыльник называют интрорзным. Если же в результате неравномерного роста связника обе половинки пыльника изгибаются и пыльца высыпается наружу, пыльник называют экстрорзным.
Иногда пыльник вскрывается почти посередине поперечной щелью, как у северо-американской пиксидантеры, у паслена, вереска, черники - двумя дырочками на верхушке, а у барбариса, лавра - клапанами, откидывающимися снизу вверх (рис. 234 К-М, П).