Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
ВстречаемостьPseudomonas fluorescens иArthrobacter в разных почвах, % колоний, учтенных на питательной среде
|
|
| зона Почвы Pseudomonas Arthrobacter |
Тундра Тундровые подзолы До 80 0—5
Лесо-луговая Подзолы и дерново- > 20 0—5
подзолистые
Сухая степь Черноземы > 15 До 10
Пустынная степь Сероземы» 8 50—60
известной степени связано с уменьшением количества гумуса в нижележащих слоях почвы, но прямая корреляция здесь отсутствует. Обычно с глубиной численность микроорганизмов снижается более резко, чем уменьшается содержание гумуса. В гумусных и нейтральных почвах микробиологический профиль, как правило, все же более глубок.
При постепенном углублении в почву значительно изменяется и характер микрофлоры. В более глубоких слоях относительно больше бацилл и часто актиномицетов. Это особенно заметно в черноземах и сероземах — почвах, где отмеченных микробных группировок больше.
Как видно из приведенных данных, в составе зимогенной микрофлоры богато представлены бактерии, особенно неспорообразу-ющие формы, родовую и видовую принадлежность которых устанавливать довольно трудно. Количество этих микроорганизмов неодинаково во всех почвах. Так, можно считать доказанным, что гнилостные бактерии Pseudomonas fluorescens, являющиеся пионерами освоения органических растительных остатков, богато представлены в почвах севера, где медленно идет процесс минерализации. В почвах юга они обнаруживаются в значительном числе лишь в течение краткого времени после внесения растительных остатков.
Представители рода Arthrobacter, по ряду признаков родственные актиномицетам, в большем числе встречаются в почвах южной зоны (табл. 10). Они свойственны более поздним стадиям распада органического вещества и предпочитают нейтральную среду. В почвах севера очень часто находится значительное количество корине-бактерий.
Из неспорообразующих азотфиксирующих бактерий Beijerinkia распространены только в кислых субтропических почвах (латеритах и желтоземах). Представители рода Enterobacter в большом количестве находятся в лесных почвах средней полосы, а рода Spirillum — в южной зоне.
Основательнее изучены группировки спорообразующих бактерий, развитие которых связано с присутствием в почве более переработанного органического вещества. Каждому типу почв свойствен характерный набор преобладающих видов бацилл (табл. 11). Другие виды здесь могут быть, но в очень малом количестве.
В почвах с более энергичными мобилизационными процессами преобладают бациллы, использующие не только органический, но и минеральный азот (Вас. megaterium, Вас. mesentericus, Вас. sub-tilis). Наоборот, в почвах со слабо протекающими процессами минерализации органических веществ доминируют спорообразующие бактерии, для которых необходим органический азот (Вас. cereus, Вас. mycoides и др.). В этом проявляется глубокая связь физиологии микроорганизмов со свойствами среды их обитания.
В достаточной степени изучена экология спорообразующих азот-фиксирующих бацилл рода Clostridium. Некоторые из них (например, Cl. pasteurianum) в больших количествах встречаются только в северных почвах. В почвах южной зоны, по данным В. Т. Емцева, доминирует Cl. acetobutylicum.
При окультуривании почвы состав почвенной микрофлоры, в том числе бацилл, существенно меняется, появляются виды спорообразующих бактерий, свойственные более южной зоне.
Иногда с органическими удобрениями (навоз или компост) в почву вносят специфические для удобрений бациллы. Так, в разогревшемся при созревании навозе содержится весьма много зародышей Вас. mesentericus, Вас. subtilis и термофильных бактерий.
С вертикальной поясностью почв связана в основном такая же смена бациллярных форм, как и с зональными почвами. Однако почвы вертикальной поясности нельзя считать полными аналогами горизонтально-зональных, поэтому и в микрофлоре их имеются некоторые различия.
Грибы. Северные почвы, имеющие кислую реакцию, наиболее богаты грибами^ Вообще в разлагающейся растительной массе и в верхних слоях почвы их биомасса больше бактериальной. Учет массы грибного мицелия в разных почвах, проведенный Т. Г. Мирчинк, показал, что в тундре на 1 г почвы приходится 4 мг мицелия грибов, в лиственных лесах — до 1 мг, а в почвах южной зоны — 0, 4— 0, 7 мг.
В почвах южной зоны родовой и видовой состав микроскопических грибов более разнообразен. В южных почвах доминируют представители рода Aspergillus, а в северных — Penicillium. По данным Т. Г. Мирчинк, род Penicillium в северных почвах представлен 35—40 видами, а в южных — лишь 10—15. Обратная картина наблюдается для грибов рода Aspergillus: в северных почвах в небольшом числе встречаются 3—5 видов этого рода, а в южных — 15—20. Северные почвы беднее, чем южные, грибами родаРивагшт (они особенно обильно размножаются в каштановых почвах и сероземах). Некоторые виды (например, Fusarium sambicinum) свойственны только щелочным почвам.
Мукоровыми грибами богаты почвы северных районов, однако некоторые роды (Choanephora, Cunninghamella, Rhizopus) приурочены к южным почвам.
В почвах обычно встречаются грибы с темнопигментированным мицелием (Dematium, Cladosporium, Macrosporium, Alternaria и т. д.). Их экология плохо изучена, но отмечается, что представители рода Dematium более распространены в почвах с малоактивными мобилизационными процессами, то есть в основном в северной зоне, а род Alternaria чаще встречается в освоенных почвах. Выявлено, что одни виды грибов рода Mortierella (M. vanaceae, М. usabellina) распространены в кислых почвах, а другие (М. alpi-na, M. dichotoma)—в нейтральных.
В настоящее время установлены индикаторные микроскопические грибы для определенных типов почв.
^ Дрожжевая микрофлора почв и растительности разных зон изучена Н. П. Бабьевой. Приведем краткое обобщение полученных ею данных.
В тундре при большой пестроте почвенного покрова основная часть дрожжей сосредоточена на мхах и торфе. Доминантные виды дрожжей в тундровых почвах не относятся к типичным педобион-там и более характерны для живых и отмирающих частей растений. Они имеют базидиомицетовую природу (представители родов Rhodotorula, Rhodosporidium, Cryptococcus).
В лесных биогеоценозах много дрожжей имеется в подстилке. Они составляют группу, относящуюся к базидиальным грибам (виды Candida, Trichosporon и др.). В минеральных горизонтах почвы дрожжей значительно меньше. Здесь доминируют типичные педо-бионты — из аскоспоровых грибов Lipomyces starkeyi и из базидио-мицетов — виды Candida и Cryptococcus.
В степном биогеоценозе травяной опад весьма богат дрожжами. Здесь встречается до 14 видов, относящихся к родам Cryptococcus, Aureobasidium, Rhodosporidium и др. Для почвы характерно доминирование Lipomyces tetrasporium.
В биогеоценозах полупустынь и пустынь на растительности доминируют дрожжи из родов Sporodiobolus, Tilletiopsis и Sporobolo-myces, образующих баллистоспоры, рассеивающиеся токами воздуха и имеющие в жизненном цикле стадии, устойчивые к засухе, — хламидоспоры. Их численность невелика. Дрожжи приурочены не к поверхностному слою почвы, они обитают на некоторой глубине — ниже 20 см. Род Lipomyces в почве пустынь отсутствует. Доминируют криптококки.
^ Стрептомицеты (актиномицеты). Группа стрептомицетов и близких к ним организмов чрезвычайно обширна и включает до 40 родов. Экология и география не только видов, но даже отдельных родов изучена более чем поверхностно. Типичные их формы, относящиеся к аэробам и образующие мицелий, широко распространены в почве. Подобно бациллам, стрептомицеты бедно представлены в северных почвах, но в южных почвах их численность резко возрастает. Это подтверждается всеми методами исследования.
Слабый рост стрептомицетов в почвах северной зоны может быть объяснен как замедленным темпом разложения здесь органического вещества, так и слабой толерантностью их к почвенной кислотности. Северные почвы имеют обычно низкие значения рН.
Разобщенные данные о видовом составе стрептомицетов разных почв можно найти в работах Е. И. Андреюк, Л. В. Калакуцкого, Н. С. Агре, Г. М. Зеновой и других ученых. Имеющиеся сведения нередко противоречивы, но все же можно сделать некоторые предварительные выводы. Так, несомненно, что почвы южной зоны не только богаче актиномицетами, но и имеют более разнообразный их видовой состав.
Очевидно, некоторые стрептомицеты распространены чрезвычайно широко (группы albus, griseus, globisporus, violaceus и аспо-рогенные формы albus). Однако группы violaceus и аспорогенные albus богаче представлены в южных почвах. Некоторые группы ак-тиномицетов (fradia, flavus, chromogenes, rubroaurantiacus) в за7 метных количествах обнаружены в серых лесных почвах. К, югу их численность возрастает.
Создается впечатление, что группа verticillatus и аспорогенные формы flavus и chromogenes тяготеют к наиболее южным почвам. Смена состава актиномицетов в разных почвах хорошо выявляется на примере пигментированных культур. Они гораздо больше распространены в почвах, формирующихся в условиях более теплого климата.
Установлено, что стрептомицеты рода Actinomadura широко распространены повсеместно, но их видовое разнообразие значительно богаче в южных почвах.
Целлюлозоразлагающие микроорганизмы. Процесс распада клетчатки, вызываемый как бактериями, так и грибами, представляет существенный интерес для познания почвообразования. Большая часть растительных остатков состоит из целлюлозы. Изучение состава микроорганизмов, разрушающих клетчатку, проведенное нами, показало, что он существенно меняется в разных почвах. В северных почвах (тундра) этот процесс связан с деятельностью некоторых медленно растущих грибов, относящихся главным образом к родам Dematium и Penicillium. В зоне тайги в этот ценоз начинают включаться микобактерии и род Cellvibrio. В южных почвах в значительной степени грибы вытесняются как указанными бактериями, так и представителями родов Rhizophlyctis и Cytophaga. В заметных количествах здесь появляются грибы рода Chaetomium. Этот ценоз в отличие от северного разрушает клетчатку быстро. Схема смены доминирующих целлюлозоразлагающих микроорганизмов при переходе из одной почвенной зоны в другую показана на рисунке 57.
Изучение особенностей физиологии разных групп целлюлозоразлагающих микроорганизмов позволило объяснить их своеобразную экологию. Эти особенности выражаются главным образом в требовательности к источникам азотного питания. Микроорганизмы северных почв (в основном грибы) могут, хотя и медленно, расти на бедных азотными соединениями средах. Микроорганизмы южных почв, разрушающие клетчатку, нуждаются в высоком уровне азотного питания. На юге процесс минерализации азота протекает значительно энергичнее, что благоприятствует развитию микроорганизмов, более требовательных к условиям среды.
Автохтонная микрофлора почвы*. Гумусовые соединения разлагаются группировкой микроорганизмов, названной С. Н. Вино-градским автохтонной. Гумус представляет собой комплекс разных по сложности соединений, весьма стойких к воздействию микроорганизмов.
Имеются фракции гумуса, существующие в почве многие сотни лет. Некоторые же фракции гумуса (например, фульвокисло-ты) разлагаются относительно легко. Поэтому при недостаточном поступлении в почву растительных остатков содержание гумуса в ней существенно снижается, в основном за счет фульвокислот.
В последнее время выявлен ряд микроорганизмов, способных достаточно энергично минерализировать гумусовые соединения почвы. К ним прежде всего следует отнести представителей рода Nocardia, называемых иногда проактиномицетами. Этот род имеет
*  От лат. antochthonous — коренная, местная.
От лат. antochthonous — коренная, местная.
лившиеся гидроокиси Fe и А1 могут вступать в реакции с фульво--кислотами и способствовать их выпадению из раствора и закреплению в аллювиальном горизонте почвы.
При избыточном увлажнении в глубине почвы создается анаэробиоз, бактерии восстанавливают окисное железо и образуются ог-леенные горизонты.
Олиготрофные микроорганизмы составляют большую группу почвенного микронаселения. Многие представители этой группы не растут на обычных питательных средах, так как не выносят высокой концентрации органических веществ. Они предпочитают ассимилировать питательные вещества из растворов с низкой концентрацией как азотсодержащих (олигонитрофилы), так и органических (олигокарбофилы) соединений.
Олиготрофы завершают минерализацию органических соединений, то есть являются группировкой, метаболически связанной с типичными представителями зимогенной микрофлоры. Олиготрофы относятся к группировке, названной Г. А. Заварзиным «микрофлорой рассеяния». Эта группировка очень разнообразна. В нее входят многие типичные сапрофиты, способные развиваться на бедных субстратах, а также ряд весьма специфических видов, отмеченных ниже. Олиготрофы могут быть учтены посевом почвенных суспензий на бедные питательные среды.
Убедиться в существовании значительной группы олиготрофов особенно в почве, удобряемой органическими веществами, можно на основании опыта, проведенного в ТСХА. В почве, паровавшей длительное время (более 50 лет), было очень мало олиготрофов (микробов, растущих на бедной среде). В почве, занятой бессменной рожью, то есть ежегодно обогащаемой пожнивными остатками, численность как зимогенной, так и олиготрофной группировки резко возрастала (рис. 59). В почве, обогащенной органикой олиго-
трофы получали хороший запас питательных веществ после разрушения пожнивных остатков зимогенной микрофлорой.
Д. И. Никитин установил, что относительная численность олиго-трофов в северных почвах намного ниже, чем в южных. Это может быть объяснено тем, что при более холодном климате минерализация органических соединений проходит медленно. В условиях же юга при достаточном увлажнении жизнедеятельность микрофлоры протекает весьма энергично. Органические остатки быстро разрушаются, и олиготрофы здесь активно размножаются.
Значительная часть олиготрофных бактерий отличается необычными морфологией и циклом развития. Они составляют большую группу так называемых новых форм микроорганизмов. Д. И. Никитин схематично делит их на следующие группы.
Почкующиеся бактерии — мелкие палочковидные микробы, обычно образующие протоплазматические гифы, на концах которых формируются почки. После созревания последние отделяются, и из них возникает новый организм. Молодые почки подвижны.
К этой группе относятся: Hyphomicrobium, имеющий неветвя-щиеся гифы; Pedomicrobium — бактерия с ветвящимися гифами; Hyphomonas, которому свойствен плеоморфизм; Blastobacter — бактерия, не образующая гиф и формирующая почки на клетке.
Простекобактерии имеют на клетке выросты (простеки) 0, 3 мкм в диаметре и менее. Эти выросты представляют собой часть клетки, включая клеточную стенку, цитоплазматическую мембрану и цитоплазму.
К простекобактериям относится ряд родов: Prosthecomicro-bium — палочковидная, размножающаяся делением бактерия, имеющая многочисленные заостренные выросты до 2 мкм в длину; Ап-calomicrobium — микроб, близкий по морфологии к предыдущему, но с более длинными (более 2 мкм) пальцевидными выростами, размножающийся почкованием; Labrys — бактерия с радиально-лучевым строением клеток, напоминающая двухлезвийный топор (греч. лабрис) и размножающаяся почкованием; Stella — бактерия, имеющая форму морской звезды с радиально-лучевой симметрией, размножающаяся делением, а также ряд других микроорганизмов.
Стебельковые бактерии имеют вибриоидные или палочковидные клетки, снабженные стебельком, который окружен общей оболочкой с клеткой. До формирования стебелька молодые клетки имеют жгутики. Стебелек служит для прикрепления к субстрату. В чистых культурах, где плотность популяций высока, многие клетки прикрепляются друг к другу, образуя розетки. Размножаются делением перетяжкой.
В группу стебельковых бактерий входят часто встречающиеся в почве Caulobacter и Asticcacaulis. У первой бактерии стебелек отходит от конца клетки, у второй — прикреплен к боковой стороне клетки.
Тороидальные, или кольчатые, бактерии. Клетки их отличаются изогнутой формой. Бактерии неподвижны. Размножаются делением. К этой группе относятся: Microcyclus, имеющий изогнутую клетку;
^ Renobacter — бактерия с клеткой палочковидной формы и Spiro-soma — со спиралевидной клеткой.
Для некоторых представителей олиготрофных микроорганизмов пока трудно определить систематическое положение. Это, в частности, относится к Seliberia — бактерии с длинными палочковидными спирально изогнутыми клетками, часто соединенными в звездообразные комплексы (розетки). Seliberia размножается делением, образуя почки. Молодые клетки Seliberia подвижны. Впервые организм был описан Т. В. Аристовской.
Совершенно очевидно, что отдельные виды олиготрофов участвуют в разложении органических соединений на разных этапах. Их экология и роль в почвенных процессах еще недостаточно изучена. ■ Ряд представителей олиготрофной группировки бактерий показан на рисунке 60.
Хемоавтотрофные микроорганизмы почвы весьма разнообразны. Они вызывают окисление неорганических соединений, образующихся при микробной трансформации преимущественно органических веществ. Наиболее изучены из этих микроорганизмов нитрифицирующие бактерии, деятельность которых характеризует энергию мобилизационных процессов в почве. Еще в 20-х годах текущего столетия сотрудниками С. П. Костычева было установлено, что по мере движения от севера к югу активность нитрификационного процесса усиливается. Это результа т усиления в условиях бол ее^теп-лого климата процессов распада органических.^сл^тков^_Г1ри которых выделяется NH3, служащий энергетическим и питательным субстратом для нитрификаторов. В условиях вертикальной зональности наблюдается аналогичное явление — по мере подъема в горы, где климат более суров, энергия нитрификационного процесса снижается.
^ К настоящему времени может считаться доказанным, что некоторые сапрофитные микроорганизмы вызывают процесс нитрификации.
При микробиологических процессах в почве образуется ряд химических веществ (Н2, H2S, CO, CH4, C2H4 и т. д.), которые могут служить источником жизни для автотрофных микроорганизмов.
^ Помимо хемоавтотрофных микроорганизмов, в почвах находятся фотоавтотрофные микроскопические существа, среди которых имеются типичные бактерии и цианобактерии.
Из отмеченных микроорганизмов в экологическом аспекте более детально изучены водоросли. В почвах арктических пустынь и тундры, на поверхности и в глубине, развиваются зеленые и желто-зеленые водоросли, много азотфиксирующих цианобактерии.
В подзолистых почвах преобладают одноклеточные зеленые (виды Chlamydomonas, Coccomyxa, Chlorococcum, Chlorella) и некоторые нитчатые зеленые. Им сопутствуют нитчатые желто-зеленые и некоторые.диатомовые водоросли. Цианобактерии, особенно в хвойных лесах, играют небольшую роль.
При дерновом процессе отмечается обильное разрастание водорослей, среди которых появляется значительное число видов циа
нобактерий, в том числе азотфиксаторов (виды Nostoc, Calothrix, Anabaena, Tolypothrix и др.)- Богато представлены зеленые и желто-зеленые водоросли.
В луговых и ковыльных степях черноземной зоны под густым травостоем водоросли развиваются менее интенсивно. Альгофлора представлена широко распространенными видами зеленых водорослей. Здесь интенсивно размножаются и цианобактерии.
В южных — сухих и полупустынных степях при разреженном травостое развитие водорослей усиливается. На поверхности каш тановых почв образуются пленки водорослей, в которых ведущая роль принадлежит цианобактериям, в том числе и азотфиксаторам. При пустынном почвообразовании состав альгофлоры напоминает таковой в полупустынной зоне, но численность водорослей существенно снижается. Преобладают цианобактерии (осциляторие-вые), распространены зеленые водоросли. Максимальное количество водорослей, по данным Э. А. Штина, Е. М. Панкратовой, наблюдается не в поверхностном слое, а на некоторой глубине.
^ На рисунке 61 дана схема структуры микробного ценоза, которая иллюстрирует взаимоотношения описанных группировок почвенных микроорганизмов.
Приведенный материал позволяет заключить, что в определенных почвах те или иные микроорганизмы отличаются различной выживаемостью, что связано с сукцессией групп микроорганизмов при трансформации минеральных и органических веществ.
В последнее время начал разрабатываться вопрос об «экологической стратегии» микроорганизмов. В это понятие вкладывают установление совокупности приспособлений, обеспечивающих виду возможность обитать совместно с другими микроорганизмами и занимать определенное положение в соответствующем биоценозе.
^ Принцип выделения «стратегов» микроорганизмов основывается на анализе динамики популяции. Могут быть намечены следующие типы стратегов:
г 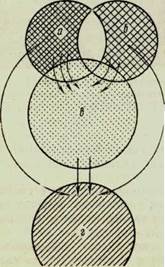 -стратеги — популяции их подвержены резким колебаниям численности. В эту группу входят зимогенные микроорганизмы, размножение которых зависит от периодических поступлений в почву органических веществ;
-стратеги — популяции их подвержены резким колебаниям численности. В эту группу входят зимогенные микроорганизмы, размножение которых зависит от периодических поступлений в почву органических веществ;
К-стратеги — устойчивые равновесные популяции, стремящиеся сохранить высокий уровень популяцион-ной плотности в имеющихся условиях. К этой группе могут быть отнесены автохтонные, олиготрофные и некото-| рые другие микроорганизмы, довольствующиеся более скромным запасом питательных веществ;
^ L-стратеги, популяции которых нередко находятся на грани вымирания, но сохраняются благодаря наличию спор.
Рис. 61. Схема структуры микробного ценоза почвы:
а — зимогенная микрофлора; б — микрофлора, разлагающая гумус; в — олиготрофные микроорганизмы; г — хемоавтотрофы.
Описанная специфика живого микронаселения почвы подтверждает закон зональности В. В. Докучаева и делает возможной микробиологическую диагностику направленности почвообразовательного процесса, плодородия почвы и его изменения под влиянием деятельности человека.
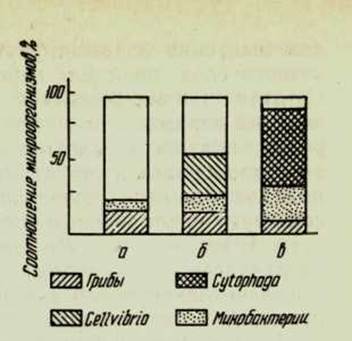
^ Рис. 62. Состав целлюлозораз-лагающих микроорганизмов в различно удобренных дерново-подзолистых почвах ТСХА:
а — контроль; б — минеральные
удобрения; в — навоз.
Некоторые группировки микроорганизмов остаются довольно константными при антропогенном воздействии на почву. Они отражают тип почвообразовательного процесса и могут быть индикаторами, которые достаточно консервативны и хранят информацию о былых состояниях факторов почвообразования. К таким сравнительно стабильным показателям относится соотношение основных групп микроорганизмов.
НИЗМОВ.
При окультуривании в микробном ценозе отмечаются существенные изменения. Увеличивается численность микробного населения, и в ценозе появляются организмы, свойственные более южной почвенной зоне. Об этом свидетельствует пример с целлюлозоразлага-ющими микроорганизмами, состав которых резко меняется при внесении удобрений (рис. 62).
^ Следовательно, микроорганизмы можно использовать для анализа типа и состояния почвы.