Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Каскадный механизм мобилизации и синтеза гликогена
|
|
Фн
Среднее содержание гликогена в печени колеблется в пределах 5-7%
а) при нормальном смешанном питании глк превращается в
- гликоген 3%
- жиры 30%
- окисляется до СО2, Н2О – 70%
б) при обильном углеводном питании глк превращается в
- гликоген 10%
- жиры 40%
- окисляется до СО2, Н2О – 50%
в) при голодании в течении суток почти весь гликоген без остатка утилизируется и его не удается обнаружить в печени обычными биохимическими реакциями.
Содержание гликогена в печени зависит от его синтеза (гликогенеза) или распада (гликогенолиза).
Распад гликогена (гликогенолиз)
происходит двумя путями
гликоген
_____________________________________________________________
Фосфоролиз (+Фн) гидролиз (+Н2О)
Глюкоза-1-фосфат
глюкоза
Глюкоза-6-фосфат
Лактат
(гликолиз)
Гидроли з гликогена (крахмала) происходит в ЖКТ под действием специфических амилаз.
Энергетически более выгодным является второй путь расщепления гликогена – фосфоролиз. Фосфоролитический распад играет ключевую роль в мобилизации полисахаридов.
(C6H10O5)n + H3PO4 гликогенфосфорилаза (C6H10O5)n–1 + Глюкозо-1-фосфат,
где (С6Н10О5)n означает полисахаридную цепь гликогена, а (С6Н10О5)n ту же цепь, но укороченную на один глюкозный остаток.
В этой реакции от нередуцирующего конца отщепляется один остаток глюкозы и соединяется в фосфорной кислотой, процесс повторяется многократно, пока весь полимер не будет расщеплен до точки ветвления. Образуется остаточный декстрин. Другой фермент изоамилаза, разрывает 1→ 6 гликозидные сязи, далее для действия фосфорилазы отрывается новый участок полисахаридной цепи. Фосфоролиз энергетически более выгоден, т.к. в результате образуется глюкоза уже в активированном состоянии – глюкоза-1-фосфат, которая легко вступает в различные реакции.
Гликоген
Фн 1
Глюкоза-1-фоосфат
Глюкоза-6-фосфат
Анаэробный гликолиз Синтез глюкозы
(лактат)
Синтез глюкозы из глицерина
1. глк-1-ф фосфоглюкомутаза глк-6-ф
2. глк-6-ф + Н2О глк-6-фосфотазапечени глк + H3PO4
Механизм фосфорилитического отщепления
остатка глюкозы от гликогена.
Гликогенфосфорилаза или просто фосфорилаза существует в двух формах: А и В.
Форма А более активна, чем В. Отличаются они друг от друга тем, что:
фосфорилаза А это тетрамер, состоящий из 4-х субъединиц с Мr=360000Да
фосфорилаза В – димер, содержащий две субъединицы, его Мr=180000Да.
Превращение неактивной формы в активную протекает по схеме:
2 фосфорилазы В + 4 АТФ акт.киназа фосфорилазы Вфосфорилаза А + 4 АДФ
Под действием адреналина и глюкогона запускается каскад реакций, который заканчивается распадом гликогена.
Каскадный механизм мобилизации и синтеза гликогена
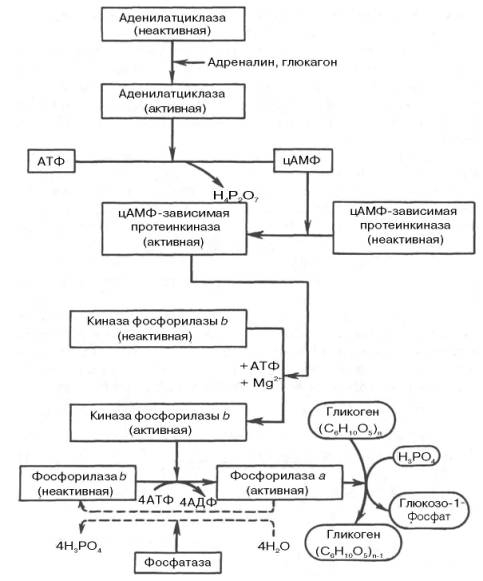
Неактивная киназа фосфорилазы превращается в активную под влиянием фермента цАМФ-зависимой протеинкиназы.
Активная форма последней образуется при участии цАМФ, которая в свою очередь образуется из АТФ под действием фермента аденилатциклазы, стимулируемой, в частности, адреналином и глюкагоном. Увеличение содержания адреналина в крови приводит в этой сложной цепи реакций к превращению фосфорилазы В в фосфорилазу А и, следовательно, к освобождению глюкозы в виде глюкозо-1-фосфата из запасного полисахарида гликогена. Обратное превращение фосфорилазы А в фосфорилазуВ катализируется ферментом фосфатазой (эта реакция практически необратима).
При отсутствии потребности организма в дополнительной глюкозе распад гликогена прекращается.
Под действием специфических ферментов происходит инактивирование фосфорилазы А и активирование гликогенсинтазы I, начинается синтез гликогена.
Т.о. активирование 1-го фермента каскада – аденилатциклазы приводит к увеличению распада гликогена и подавлению его синтеза.
Гликолиз
В клетках организма человека и других высших организмов образование энергии происходит не только в просе окислительного фосфорилирования (аэробным путем), но и в ходе распада питательных веществ без участия О2 (анаэробным путем). Главным источником получения энергии анаэробным путем служат гексозы, прежде всего α -Д-глюкоза.
Гликолиз – это анаэробный распад глк до 2-х молекул молочной кислоты (лактата), протекающих в гиалоплазме клетки.
Суммарное уравнение анаэробного гликолиза

Это экзергонический процесс (выделяется энергия). Освобождающаяся в результате гликолиза энергия аккомулируется в фосфатных связях АТФ.
По имени авторов, изучавших этот процесс более проблемно, гликолиз называют путем Элебдена-Мейергофа-Парнаса.
Отдельные стадии гликолиза катализируются 11-ю ферментами.
Анаэробный гликолиз можно подразделить на две стадии:
| 1 стадия - | активирование глюкозы и дальнейший ее распад на две триозы (5-я р-ция) |
| 2 стадия - | Включает окислительно-восстановительную реакцию (р-цию оксиредукции) сопряженную с субстратным фосфорилированием, в процессе которого образуется АТФ |
СТАДИЯ
1.↓ Р-ция фосфорилирования глк, т.е. перенос остатка ортофосфата на глюкозу за счет АТФ. Реакция катализируется ферментом гексокиназой:

Образование глюкозо-6-фосфата в гексокиназной реакции сопровождается освобождением значительного количества свободной энергии системы и процесс практически необратим. Глк-6-ф в отличии от глк, не проходит через мембрану и как бы «запирается в клетке».
Гексокиназа существует в виде четырех изомеров. Первые три преобразуют различные виды гексоз, включая глк. Гексокиназа IV (глюкокиназа) фосфорилирует только глк, имеется в печени, включается в работу при значительных концентрациях глк в крови воротной вены. Остальные изоформы существуют во всех органах и тканях. Гексокиназа ингибируется глюкозо-6-фосфатом, т.е. последний служит одновременно и продуктом реакции, и аллостерическим ингибитором.
2.↓ ↑ Изомеризация глюкозо-6-фосфата в фруктозо-6-фосфат:

Эта реакция протекает легко в обоих направлениях, и для нее не требуется каких-либо кофакторов.
3.↓ Образовавшийся фруктозо-6-фосфат фосфорилируется за счет второй молекулы АТФ:
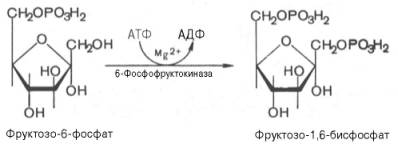
Данная реакция аналогично гексокиназной практически необратима и является наиболее медленно текущей реакцией гликолиза, так как здесь происходит резкое падение свободной энергии.
Фермент фосфофруктокиназа по своей природе аллостерический фермент. Она ингибируется АТФ и стимулируется АМФ. При значительных величинах отношения АТФ/АМФ активность фосфофруктокиназы угнетается и гликолиз замедляется. Напротив, при снижении этого коэффициента интенсивность гликолиза повышается. Во время работы мышцы происходит интенсивное потребление АТФ и активность фосфофруктокиназы повышается, что приводит к усилению процесса гликолиза.
Фосфофруктокиназа – это «ключевой» фермент гликолиза, он лимитирует скорость всего процесса гликолиза.
4.↓ ↑ Под влиянием фермента альдолаза фруктозо-1, 6-дисфосфат расщепляется на две фосфотриозы:
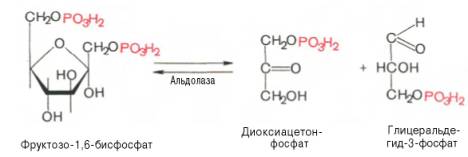
Эта реакция обратима. При повышении температуры реакция сдвигается в сторону большего образования триозофосфатов дигидроксиацетонфосфата (ДАФ) и глицеральдегид-3-фосфата (ГАФ)).
5.↓ ↑ Реакция изомеризации триозофосфатов.

Равновесие данной реакции сдвинуто в сторону дигидроксиацетонфосфата. Образуется ≈ 95% дигидрокси ацетон фосфата и 5% глицер альдегид -3-фосфата. В последующие реакции гликолиза включается только глицеральдегид-3-фосфат. По мере его потребления в ходе дальнейших превращений дигидроксиацетонфосфат превращается в глицеральдегид-3-фосфат.
СТАДИЯ
6. ↓ ↑ Окисление глицеральдегид-3-фосфата в присутствии фермента глицеральдегид-фосфатдегидрогеназы, кофермента НАД и неорганического фосфата (гликолити-ческая оксидоредукция) с образованием 1, 3-бисфосфоглицериновой кислоты и восстановленной формы НАД (НАДН2).

Реакция обратима, продукты реакции блокируют фермент, поэтому необходимо использовать их непрерывно в ходе реакции.
7. ↓ ↑ Передача богатого энергией фосфатного остатка (фосфатной группы в положении 1) на АДФ с образованием АТФ и 3-фосфоглицериновой кислоты (3-фосфоглицерата):

Р-ция экзергоническая, происходит значительное падение свободной энергии, равновесие сдвинуто влево.
Реакция обратима – это первая реакция гликолиза где образуется АТФ. В отличие от окислительного фосфорилирования образование АТФ из высокоэнергетических соединений называется субстратным фосфорилированием.
8. ↓ ↑ Реакция изомеризации. Сопровождается внутримолекулярным переносом оставшейся фосфатной группы.
Реакция легкообратима, протекает в присутствии ионов Mg2+.

9. ↓ ↑ Реакция дегидратации. Катализируется ферментом енолазой, при этом 2фосфоглицериновая кислота в результате отщепления молекулы воды переходит в фосфоенолпировиноградную кислоту (фосфоенолпируват). В результате образуется еще 1 макроэргическая связь. Реакция обратима.

Енолаза активируется двухвалентными катионами Mg2+ или Мn2+ и ингибируется фторидом.
10. ↓ 2-ая реакция гликолитического фосфорилирования. Реакция необратима, т.к. характеризуется резким падением свободной энергии, разрывом высокоэргической связи и переносом фосфатного остатка от фосфоенолпирувата на АДФ (субстратное фосфорилирование). Синтезируется еще одна молекула АТФ.

Активируется в присутствии ионов Mg2+, одновалентных катионов щелочных металлов (К+ или др.).
11. Восстановление пировиноградной кислоты до молочной кислоты. Реакция протекает при участии фермента л актатдегидрогеназы и кофермента НАДН2, образовавшегося в шестой реакции:

Продукт реакции лактат является «тупиком» в обмене веществ, т.к. не вступает ни в один биохимический процесс, кроме обратного превращения в ПВК. При накоплении лактата в клетке нарушается ее рН и останавливается гликолиз.
Т.о. Анаэробный гликолиз – это распад глюкозы до ПВК (с 1-ой до 10 р-ции). При потребности в большем количестве энергии происходит дальнейшее окисление ПВК (ее окислительное декарбоксилирование, преобразование в ацетил-КоА и полное окисление в ЦТК.
Энергетический баланс и биологическая функция гликолиза.
Распад АТФ происходит в 1-ой и 3-ей реакциях, на стадии фосфорилирования глк и фрк.
Если учесть, что весь дигидроксиацетон (5-я реакция), превращается в альдегид, то дальше происходят биохимические превращения двух триоз, а значит образование 4-х молекул АТФ в двух стадиях гликолитического фосфорилирования, т.е. образование двух молекул АТФ на одну молекулу глюкозы.
Три фермента – гексокиназа, фосфофруктокиназа, пируваткиназа (три необратимые реакции) лимитируют скорость гликолиза. Кроме того, контроль гликолиза осуществляется также ЛДГ и ее изоферментами. Воздействуя на них можно регулировать эту скорость.
Хотя анаэробный гликолиз дает малый выход энергии (2 молекулы АТФ на 1 мол глк), это единственный процесс в клетках организма, образующий энергию в отсутствии кислорода.
Поэтому в кризисных ситуациях (гипоксия) роль анаэробного гликолиза неоценимо велика, т.к. гликолиз протекает во всех клетках и тканях. В этом биологическая роль гликолиза.
Энергетическая ценность аэробного гликолиза.
При окислении 1 молекулы глюкозы в аэробных условиях (с 1-10 реакции) образуется 2 молекулы АТФ. 10-я реакция заканчивается образованием ПВК. Причем при распаде 1 молекулы глюкозы образуется 2 молекулы ПВК.
В реакциях окислительного декарбоксилирования 2-х ПВК
2 мол. НАД·H2 · 3 ═ 6 мол АТФ
В ЦТК 6 мол. НАД·H2 · 3 ═ 18 мол АТФ
2 мол. ФАД·H2 · 2 ═ 4 мол АТФ
Субстр. Фосфорилир-е = 2 мол АТФ
В гликолизе 2 мол АТФ
32 молекулы АТФ образуется при окислении 1 молекулы глюкозы в аэробных условиях.

- Регуляция гликолиза. Эффект Пастера – снижение скорости потребления глюкозы и накопления лактата в присутствии кислорода.
- Эффекта Пастера объясняется наличием конкуренции между ферментами аэробного (ПВК ДГ, ПВК карбоксилаза, ферменты цепи окислительного фосфорилирования) и анаэробного (ЛДГ) пути окисления за общий метаболит ПВК и кофермент НАДН2.
- Без О2 митохондрии не потребляют ПВК и НАДН2, в результате их концентрация в цитоплазме повышается и они идут на образование лактата. Так как анаэробный гликолиз дает из 1 глюкозы только 2 АТФ, для образования достаточного количества АТФ необходимо много глюкозы (в 19 раз больше чем в аэробных условиях).
- В присутствии О2, митохондрии выкачивают ПВК и НАДН2 из цитоплазмы, прерывая реакцию образования лактата. При аэробном окислении из 1 глюкозы образуется 38 АТФ, соответственно для образования достаточного количества АТФ необходимо мало глюкозы (в 19 раз меньше чем в анаэробных условиях).
ТЕМА: УГЛЕВОДЫ III
Цель: Дать представление о основных путях тканевых превращений глюкозы, механизмы регуляции углеводного обмена, патологии, возникающие в результате его нарушения.
ПЛАН ЛЕКЦИИ
1. Глюконеогенез, обходные пути необратимых реакций
2. Пентозофосфатный цикл окисления глюкозы.
3. Регуляция углеводного обмена.
4. Патология углеводного обмена.
Глюконеогенез – это синтез глюкозы из неуглеводных продуктов: молочной и пировиноградной кислоты, так называемых гликогенных аминокислот, глицерина и т.д.
Главное место процесса печень, в меньшей степени почки и слизистая жкт.
Глюконеогенез протекает при длительном голодании. Можно было бы считать, что глюконеогенез это процесс обратный анаэробному гликолизу. Но 3 реакции гликолиза (гексокиназная, фосфофруктокиназная и пируваткиназная) необратимы, путь обратный гликолизу «снизу вверх» невозможен, поэтому в процесс глюконеогенеза на 3 этапах существуют обходные пути необратимых реакций и используются другие ферменты.
I Обходной путь (путь синтеза глюкозы из пирувата)
Образование фосфоенолпирувата из пирувата в обход пируваткиназы

1 - пируваткарбоксилаза; 2 - малатдегидрогеназа (митохондриальная); 3 -малатдегидрогеназа (цитоплазматическая); 4 - фосфоенолпируваткарбоксикиназа.
Синтез фосфоенолпирувата осуществляется в несколько этапов.
1.(№10) Первоначально пируват проникает в митохондрии, под влиянием пируваткарбоксилазы и при участии СО2 и АТФ карбоксилируется с образованием оксалоацетата (ЩУК):

2. Затем оксалоацетат возвращается в цитоплазму и в результате декарбоксили-рования и фосфорилирования под влиянием фермента фосфоенолпируваткарбоксилазы превращается в фосфоенолпируват. Донором фосфатного остатка в реакции служит гуанозинтрифосфат (ГТФ):

Далее идут обратимые реакции гликолиза до следующей необратимой реакции. Фосфоенолпируват, образовавшийся из пирувата, в результате ряда обратимых реакций гликолиза превращается во фруктозо-1, 6-бисфосфат.
II Обходной путь (обход фосфофруктокиназной реакции (№3), которая необратима)
1. Глюконеогенез идет в обход этой эндергонической реакции. Превращение фруктозо-1, 6-дифосфата (бисфосфата) во фруктозо-6-фосфат, катализируется специфи-ческой фосфатазой:

2. В последующей обратимой стадии биосинтеза глюкозы фруктозо-6-фосфат превращается в глюкозо-6-фосфат.
фруктозо-6-фосфата глюкозофосфатизомераза глюкозо-6-фосфата
III. Обходной путь гексокиназной реакции (№1)
