Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Структура и функции гипоталамо-гипофизарно-тиреоидной системы
|
|
Глава 1 ОБЗОР ЛИТЕРАТУРЫ
Тиреоидная система организма является одной из ведущих систем, определяющих функциональное состояние организма в норме и при патологии.
Щитовидная железа(glandula thyroidea) - это непарный орган, располагающийся в передней области шеи на уровне гортани и верхнего отдела трахеи. Состоит из правой и левой доли и перешейка. Железа имеет фиброзную капсулу, от которой в глубину ткани отходят соединительнотканные перегородки — трабекулы, разделяющие железу на дольки, состоящие из фолликулов. Внутри стенка фолликулов выстлана эпителиальными клетками кубической формы. Внутри полости фолликула находится густое вещество — коллоид, которое содержит тиреоидные гормон [5].
Функция щитовидной железы (ЩЖ) регулируется преимущественно тиреотропным гормоном (ТТГ) – гликопротеином, выделяемым гипофизом. В норме регуляция тиреоидной функции во многом зависит от факторов, регулирующих синтез и секрецию ТТГ. К последним относятся тиреотропин-рилизинг-гормон (ТРГ), а также уровень циркулирующих тиреоидных гормонов, которые по принципу «обратной связи» влияют на продукцию ТТГ. Таким образом, на продукцию ТТГ, с одной стороны, оказывает стимулирующее воздействие ТРГ, а с другой – супрессивный эффект гормонов ЩЖ [6].
Результатом динамического взаимодействия этих двух доминирующих влияний на секрецию ТТГ является довольно стабильная концентрация ТТГ в циркулирующей крови. Наличие столь тонкого регуляторного механизма позволяет говорить о том, что обнаружение аномального уровня ТТГ в крови в подавляющем большинстве случаев свидетельствует о нарушении функции ЩЖ. Понимание процесса регуляции секреции ТТГ лежит в основе как нормальной физиологии ЩЖ, так и патофизиологии тиреоидных заболеваний.
Общая схема регуляции секреции ТТГ соответствует модели, приведенной на рисунке 1. Как уже отмечалось, пептидергические нейроны в преоптической области гипоталамуса синтезируют и выделяют в воротную систему гипофиза тиреотропин-рилизинг гормон (ТРГ). Определение уровня ТТГ в крови в течение суточных циклов сон — бодрствование показывает, что ТРГ, подобно другим рилизинг-факторам, эпизодически секретируется как днем, так и ночью, но пик содержания ТТГ приходится на часы, непосредственно предшествующие сну. Затем в течение ночи его уровень снижается, свидетельствуя о появлении во время сна ингибиторных влияний на секрецию ТРГ [7].
Гипоталамо-гипофизарно-тиреоидная система представляет собой функциональную суперсистему, работающую по принципу обратных связей. Основным звеном механизма обратной связи является изменение чувствительности клеток аденогипофиза к стимулирующему действию ТРГ в зависимости от концентрации тиреоидных гормонов.
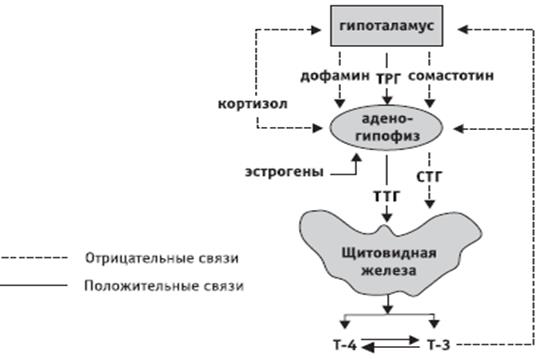
Рисунок 1. Схема регуляции гипоталамо-гипофизарно-тиреоидной системы
Уровень тиреоидных гормонов в периферических тканях определяет выработку гипоталамического тиреолиберина, который в свою очередь регулирует биосинтез и освобождение в портальную систему гипофиза тиреотропного гормона (ТТГ) [7].
Развитие гипоталамо-гипофизарного контроля функции щитовидной железы у человека происходит в период между 20 и 30-ой неделями антенатального развития и в первом месяце постнатальной жизни. В основе регуляции секреции ТТГ лежит механизм отрицательной и положительной обратной связи: высокие концентрации свободных Т4 и ТЗ ингибируют, а низкие - стимулируют его выброс. Необходимо помнить, что в аденогипофизе дейодирование Т4 с образованием ТЗ идет значительно более интенсивно, чем в периферических тканях. Поэтому, уровень ТТГ, определяемый в крови не претерпевает мгновенных изменений при назначении того или иного лекарственного препарата, а наблюдается только через некоторое время.
ТТГ представляет собой гликопротеид с молекулярной массой 28 000, состоящий из двух субъединиц — альфа и бета. Период полураспада ТТГ составляет 40-60 минут. Биологическая активность ТТГ осуществляется его бета-субъединицей. ТТГ оказывает прямое действие на щитовидную железу. Одной из причин изменения секреции тиреоидных гормонов в результате нарушения центральных регуляторных механизмов является повышенная или сниженная секреция ТТГ [8].
На поверхности мембран тиреоцитов присутствуют специфичные для альфа-субъединицы ТТГ рецепторы. Под действием ТТГ образуется циклический моноаминофосфат, запускающий каскад фосфорилирования ряда белковых субстратов, что приводит к реализации биологического эффекта ТТГ — синтезу гормонов щитовидной железы.
Принцип обратной афферентации или принцип обратных связей в гипоталамо-гипофизарно-тиреоидной системе лежит в основе исследования функционального состояния щитовидной железы в норме и при различных заболеваниях. Знание этого принципа необходимо для коррекции проводимой терапии. Например, удаление щитовидной железы или применение тиреостатических препаратов, сопровождается увеличением содержания ТТГ в крови. Соответственно этому при первичном гипотиреозе у людей наблюдается повышенный уровень ТТГ, а нормализация уровня тиреоидных гормонов сопровождается снижением ТТГ. Также, не совсем ясна роль ТТГ в возникновении нетоксического узлового зоба [9].
Долгое время считалось, что развитие зоба зависит от секреции ТТГ, однако, в последнее время было установлено, что уровень ТТГ при узловом зобе чаще всего не изменен и у больных, особенно в возрасте старше 50 лет, имеет место ТТГ-независимая реакция на тиреолиберин. Причина отсутствия реакции ТТГ на тиреолиберин при узловом эутиреоидном узловом зобе не выяснена. Можно предполагать, что эутиреоидное состояние у таких больных поддерживается секрецией ТЗ, а это влияет на состояние системы " обратной связи".
С возрастом секреторная функция щитовидной железы снижается. Возрастное уменьшение среднесуточной концентрации общего Т4 в крови и его свободной фракции у мужчин наступает раньше, чем у женщин. Вместе с тем, на введение тиреолиберина сохраняется адекватная реакция щитовидной железы, что свидетельствует об интактности гипоталамо-гипофизарно-тиреоидных связей, а также достаточности функциональных резервов железы.
Контроль регуляции тиреоидной функции осуществляется и на уровне щитовидной железы. Йодная недостаточность приводит к гиперсекреции ТТГ, а тиреоидные гормоны могут угнетать функцию щитовидной железы независимо от гипоталамуса и гипофиза. Помимо центральных, гипоталамо-гипофизарных механизмов регуляции функции щитовидной железы, существует периферическая регуляторная система, влияющая на секрецию тиреоидных гормонов. Основная роль в этой системе принадлежит тиреостимулирующим иммуноглобулинам. Действие иммуноглобулинов заключается в увеличении поглощения йода щитовидной железой, ускорением высвобождения тиреоидных гормонов и индукции гистологических изменений в ткани щитовидной железы, неотличимых от действия ТТГ [10].
Основными и необходимыми компонентами синтеза тиреоидных гормонов — тироксина (Т4) и трийодтиронина (ТЗ) являются йод, поступающий в адекватных количествах в щитовидную железу, и аминокислота тирозин. Йод поступает в организм с пищевыми продуктами. Физиологическое потребление йода человеком составляет 150-200 мкг в сутки. Всосавшийся из кишечника йод в виде йодидов достигает посредством кровеносной системы щитовидной железы и активно проникает через базальную мембрану в фолликулярные клетки против градиента концентрации. Йодид претерпевает стадию окисления, в результате чего переходит в молекулярный йод. Молекулярный йод соединяется с тиреоглобулином и в свободной форме остается всего 1-2% йода.
На рисунке 2 показано строение некоторых биологически важных иодированных соединений.
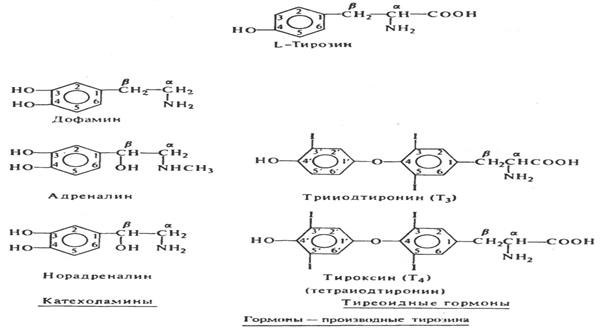
Рисунок 2. Тирозин и некоторые его производные
Органификация йода происходит в тиреоцитах, куда тиреоглобулин проникает через коллоид. Именно там осуществляется органическое связывание йода с последовательным образованием монойодтирозина (МИТ) и дийодтирозина (ДИТ).
В результате окислительной конденсации двух молекул ДИТ с потерей одной аланиновой цепи образуется тироксин. Образование трийодтиронина происходит в результате соединения молекул ДИТ и МИТ также с потерей одной аланиновой цепи.
Секреция тиреоидных гормонов начинается с резорбции коллоида под влиянием протеолитических ферментов. В результате протеолиза освобождаются МИТ, ДИТ, Т4 и ТЗ. МИТ и ДИТ подвергаются обратному дейодированию и высвобождающийся в результате этого йод вновь используется в синтезе тиреоидных гормонов.
В кровоток в основном поступают ТЗ и Т4 и циркулируют там в связанной транспортными белками форме. Щитовидная железа секретирует в 10-20 раз больше Т4 чем ТЗ, однако, ТЗ активнее Т4 по своему действию в 5 раз. Период полувыведения Т4 из организма составляет 6-7 дней, причем, около 40% тироксина метаболизируется с образованием ТЗ и реверсивного (неактивного) ТЗ. Период полураспада ТЗ равен 1-2 дням.
На периферии дейодированию подвергаются как Т4 так и ТЗ собразованием тетрайодтиропропионовой, тетрайодтироуксусной и трийодтироуксусной кислот. Эти вещества обладают очень слабым метаболическим эффектом.
Хотя природа фактора, ингибирующего секрецию тиреотропина, окончательно не выяснена, некоторые данные указывают на то, что эту физиологическую функцию может выполнять соматостатин. Показано, что у крыс, пассивно иммунизированных антисывороткой к соматостатину, повышается как базальный уровень ТТГ, так и его реакция на введение ТРГ. Это свидетельствует о возможности тонического контроля секреции ТРГ со стороны соматостатина. Кроме того, ингибирование секреции ТТГ дофамином и повышение ее после введения метоклопрамида (блокирующего эффекты дофамина) указывают на физиологическую роль дофамина как фактора, ингибирующего секрецию ТТГ. Подобно тому, что происходит при торможении секреции пролактина, дофамин, вероятно, непосредственно влияет на клетки гипофиза [11].
Основное ингибирующее действие на секрецию ТТГ оказывает повышенная концентрация тиреоидных гормонов в крови. Механизм этого действия состоит в том, что спустя определенный лаг-период, в течение которого может происходить синтез белка, гипофизарные тиреотрофы приобретают резистентность к стимулирующему действию ТРГ. В настоящее время счита-ют, что тормозящее действие тиреоидных гормонов по принципу обратной связи осуществляется в основном на гипофизарном уровне, хотя обнаружение связывающих участков для трииод-тиронина (Т3) в гипоталамусе не позволяет отрицать регуляторного влияния тиреоидных гормонов и на секрецию ТРГ.
Реакция гипофиза на ТРГ модулируется и другими гормонами. Эстрогены повышают чувствительность тиреотрофов к ТРГ и усиливают секреторную реакцию ТТГ на этот фактор.
Глюкокортикоиды и гормон роста ингибируют реакцию тиреотрофов на ТРГ. Моноаминергическая нервная регуляция ТРГ-секретирующих нейронов изучена не столь подробно, как в отношении других пептидергических нейронов, но известные супрагипоталамические факторы, влияющие на секрецию ТРГ — сон, низкая температура и неспецифический стресс — действуют, по-видимому, через посредство различных нейронных цепей и с помощью разных нейромедиаторов. Наиболее важным из медиаторов, стимулирующим секрецию ТРГ, является, вероятно, норадреналин, действующий через α -рецепторы.
Как уже отмечалось, ТРГ - мощный стимулятор секреции не только ТТГ, но и пролактина. Действительно, его применяют для усиления секреции молока, если у женщины ослаблена реакция на грудное вскармливание ребенка. В то же время ТРГ не может быть ПРЛ-рилизинг-фактором, поскольку существуют условия, при которых секреция ТТГ и ПРЛ происходит по-разному.
Открытие ТРГ — первого очищенного, идентифицированного и синтезированного in vitro гипоталамического фактора — было событием в биологии. Последующая разработка высокочувствительных методов определения этого трипептида позволила установить, что, хотя концентрация ТРГ в гипоталамусе весьма велика, более 80% его содержится во внегипоталамическнх отделах мозга.
Большие количества ТРГ обнаружены в эпифизе. По-видимому, внегипоталамический ТРГ синтезируется именно там, где обнаруживается; во всяком случае, он сохраняется в других отделах мозга после разрушения ТРГ-продуцирующих нейронов гипоталамуса.
Стимуляция секреции ТТГ на первых порах не сопряжена с активацией синтеза белка, но длительное воздействие ТРГ в конце концов каким-то образом приводит к повышению синтеза ТТГ (рисунок 3).
Функция ТРГ вне гипоталамуса неизвестна; сообщалось о влиянии этого трипептида на поведенческие реакции и электрофнзиологические процессы в нервной системе. Механизм действия ТРГ на тиреотрофы включает связывание его со специфическими рецепторами и стимуляцию секреции ТТГ путем увеличения концентрации Са2+ в цитозоле.
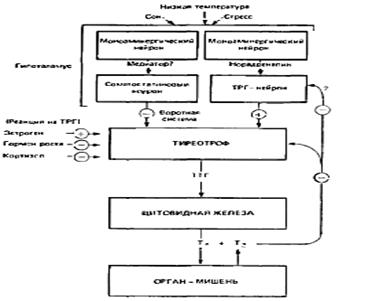
Рисунок 3. Иерархическая схема регуляции секреции
тиреодных гормонов
Вероятно, в качестве вторых посредников в этом процессе участвуют продукты превращения полифосфатидилинозитола, судя по тому, что добавление ТРГ к клеткам гипофиза (линия GH3) приводит к повышению в них концентрации диацилглицерола. Правда, до сих пор не доказано, что тиреотрофы реагируют на ТРГ точно так же, как клетки GH3 [12].