Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
История изучения нуклеиновых кислот. Доказательства генетической роли нуклеиновой кислоты
|
|
История изучения нуклеиновых кислот начинается с 1869 г., когда швейцарский химик И.Ф. Мишер обнаружил в клеточном ядре особые вещества, обладающие свойствами кислот. Он дал им название нуклеиновых кислот (от лат. nukleus — ядро) (рис. 5.1).

Рис. 5.1. Мишер Иоганн Фридрих (1844-1895)
В 1889 г. немецкий гистолог Рихард Альтман обозначил кислый компонент нуклеина термином «нуклеиновая кислота». В конце XIX в. немецкий биохимик Альбрехт Коссель (1853-1927) (рис. 5.2) расшифровал химический состав нуклеиновой кислоты, показав, что она содержит фосфорную кислоту, углевод и азотистые основания (пурины и пиримидины).
Ф. Левен, Д. Гулланд с сотрудниками (в 1900-1932 гг.) установили, что фосфорная кислота, углевод и азотистое основание соединены в блоки в виде мономеров – нуклеотидов.
Нуклеиновая кислота, выделенная из ядер клеток, в качестве углевода содержит D-дезоксирибозу. Поэтому она получила название дезоксирибонуклеиновой кислоты – ДНК. Наряду с ядерной была выделена цитоплазматическая нуклеиновая кислота, содержащая в качестве углевода D-рибозу; она получила название рибонуклеиновой кислоты – РНК.

Рис. 5.2. Альбрехт Коссель (1853-1927)
А.Н. Белозерский и И.И. Дубровская в 1936 г. выделили ДНК из ростков конского каштана. Это показало, что ДНК входит в состав клеток и животных и растений. ДНК была обнаружена и в клетках бактерий.
Современные взгляды на химическое строение нуклеиновых кислот сформировались в 40-50-х гг. прошедшего века.
Впервые прямые доказательства того, что молекулы ДНК являются носителями наследственности, были получены при исследовании у бактерий явления трансформации и позже подтверждены результатами исследования трансдукции. Вкратце эти исследования состояли в следующем.
Долгое время нуклеиновые кислоты не привлекали внимания исследователей. И только после того как в опытах английского бактериолога Ф. Гриффита (1928) была продемонстрирована способность пневмококков к трансформации, было выдвинуто предположение о том, что «трансформирующий агент», отождествляемый с «веществом наследственности», находится в ядре.
Суть эксперимента Гриффита заключалась в следующем. При введении мышам непатогенных штаммов пневмококков (рис. 5.3) животные не заболевали. При введении патогенных штаммов мыши гибли, однако при введении патогенных микробов, убитых нагреванием, мыши оставались здоровыми.
Гриффит показал, что при одновременном введении живых непатогенных и убитых патогенных микробов мыши погибали. Гриффит заключил, что живые микробы непатогенного штамма в присутствии клеток штамма патогенного приобретают наследственно закрепленные свойства патогенности (трансформируются).
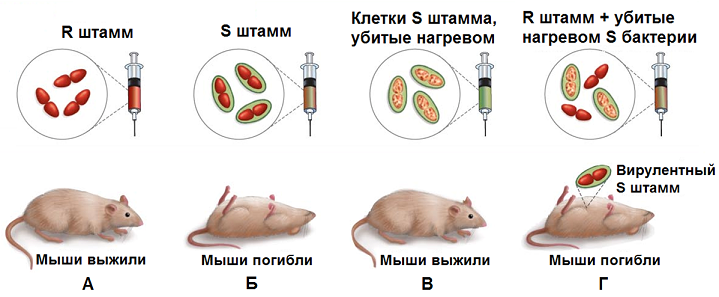
Рис. 5.3. Эксперимент Ф. Гриффита
В 1944 г. химическая природа трансформирующего агента у пневмококков была изучена О. Эйвери, К. Мак-Леодом и М. Мак-Карти. Они установили, что полисахариды из капсулы пневмококков, белки из их клеток, а также рибонуклеиновые кислоты (РНК) трансформирующим эффектом не обладали. И только молекулы ДНК из капсульных бактерий были способны вызвать трансформацию.
Трансформирующая способность ДНК была подтверждена специальным экспериментом. Фермент дезоксирибонуклеаза разрушает молекулы ДНК. После обработки ДНК дезоксирибонуклеазой, трансформирующая фракция теряла способность вызывать трансформацию.
В 1952 г. А.Херши и М.Чейз проводили эксперименты с бактериофагом Т2 — особым видом вируса, убивающим зараженную бактериальную клетку (рис. 5.4).
Бактериофаг, проникая в кишечную палочку Еscherichia coli (Е. соli), быстро в ней размножается. Экспериментаторы размножали бактериофаги в клетках Е. соli, которые росли на двух различных средах: на среде, содержащей радиоактивный изотоп серы (35S), и на среде, содержащей радиоактивный изотоп фосфора (32Р).
Фаги, которые размножились на клетках, выросших на среде с радиоактивным изотопом серы, включали 35S только в свои белковые оболочки. Фаги, размножившиеся на клетках, которые выросли на среде с радиоактивным фосфором, содержали ДНК, меченную 32Р. Затем полученными бактериофагами заражали клетки Е. соli, выращенные на обычной среде.
Оказалось, что бактерии, зараженные фагами, выросшими на 35S, не содержали радиоактивной метки, в то время как бактерии, зараженные фагами, размножившимися на 32Р, были радиоактивными.
Полученные результаты позволили авторам сделать два принципиальных вывода:
1) в бактериальную клетку проникает только фаговая ДНК, которая, размножаясь в клетке Е. соli, дает начало многочисленному потомству;
2) наследственным материалом является ДНК, которая определяет не только структуру и свойства ДНК потомства, но и свойства фаговых белков.
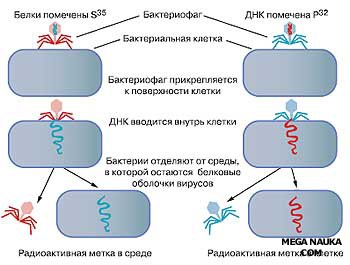
Рис. 5.4. Эксперимент А.Херши и М.Чейз
Позже, в 1956 г. было показано, что у РНК-содержащих вирусов генетическая информация также представлена нуклеиновой кислотой, а не белком. Доказано это было в экспериментах с вирусом табачной мозаики.