Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Пространственная модель ДНК
|
|
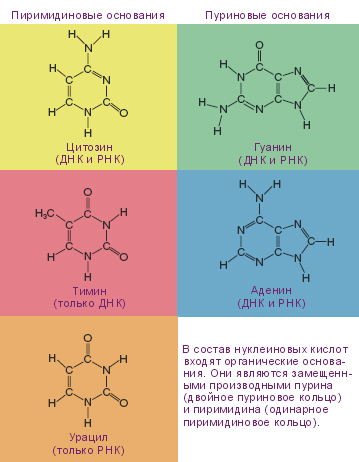
Рис. 5. Азотистые основания, входящие в состав нуклеиновых кислот
Американский биохимик Эрвин Чаргафф разработал точные методы определения количества азотистых оснований и установил характерные особенности химического состава нуклеиновых кислот. Это сыграло большую роль в познании молекулярной структуры ДНК. Им было установлено, что азотистые основания, входящие в состав ДНК (рис. 5.5) и выделенные из клеток различных организмов (рис. 5.8) подчиняются закономерностям. Сумма пуриновых оснований (А + Г) всегда равна сумме пиримидиновых (Ц + Т).
Содержание аденина равно содержанию тимина, а содержание гуанина — количеству цитозина A=Т; Г=Ц (рис. 5.6). Данные правила предложил ученый Эрвин Чаргафф (рис. 5.7).

Рис. 5.6. Правило Чаргаффа

Рис. 5.7. Эрвин Чаргафф (1905-2002)
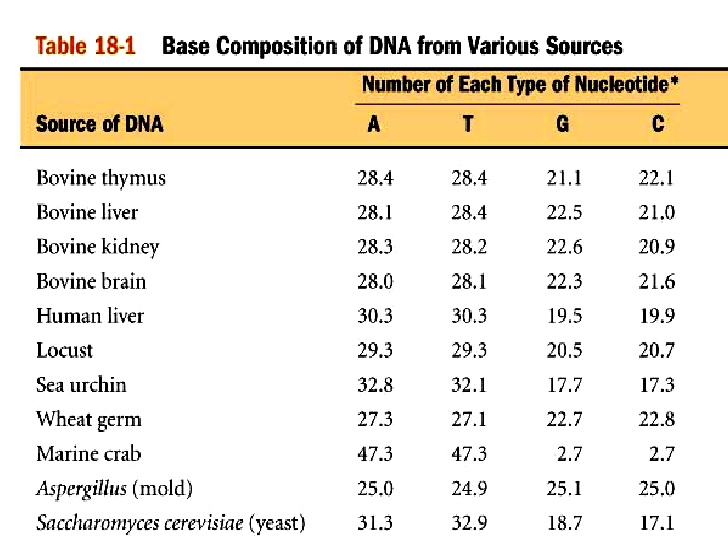
Рис. 5.8. Азотистые основания, выделенные из клеток различных организмов
В 1953 г. американский молекулярный биолог Джеймс Уотсон и английский физик и генетик Френсис Крик (рис. 5.9), основываясь на данных Э. Чаргаффа и М. Уилкинса, а также Розалинда Франклин (рис. 5.10) построили модель пространственной структуры молекулы ДНК. Это открытие было удостоено высшей научной награды — Нобелевской премии.

Рис. 5.9. Джеймс Уотсон и Френсис Крик (1953)

Рис. 5.10. Розалинда Франклин (1920-1958), английский биофизик и учёный-рентгенограф
В соответствии с моделью Дж. Уотсона и Ф. Крика молекула ДНК состоит из двух длинных комплементарных полинуклеотидных цепей, закрученных в правильную двойную спираль.
Диаметр двойной правозакрученной спирали ДНК составляет около 2 нм, один поворот спирали (шаг) – 3, 4 нм. В каждом витке (шаге) спирали находится 10 пар нуклеотидов, расстояние между нуклеотидами равно 0, 34 нм (рис. 5.11).

Рис. 5.11. Третичная структура ДНК
Скелетная основа полинуклеотидных цепей содержит правильно чередующиеся сахара и фосфаты, связанные ковалентными связями. Две углеводно-фосфатные цепи расположены на внешней стороне молекулы ДНК, в то время как азотистые основания находятся внутри ее, перпендикулярно оси спирали.
Аденин одной цепи соединяется двумя водородными связями с тимином другой цепи.
Между гуанином и цитозином образуются три водородные связи.
Такое соединение азотистых оснований обеспечивает прочную связь двух цепей и сохранение равного расстояния между ними на всем протяжении и называется комплементарностью (рис. 5.14).
Комплементарность – это пространственная взаимодополняемость молекул или их частей, приводящая к образованию водородных связей.
Комплементарность каждой отдельной пары оснований создаёт комплементарность двух полинуклеотидных цепей в целом.
Водородные связи возникают между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи в результате избирательного спаривания оснований.
Соединение одного из пуринов (А или Г) или пиримидинов (Ц или Т) с остатком сахара образует нуклеозид.
После присоединения к нуклеозиду фосфатной группы возникает нуклеотид, содержащий основание, сахар и фосфатную группу. Фосфатная группа присоединяется к нуклеозиду, заменяя в дезоксирибозе группу ОН– в положении 5′ (рис. 5.12).
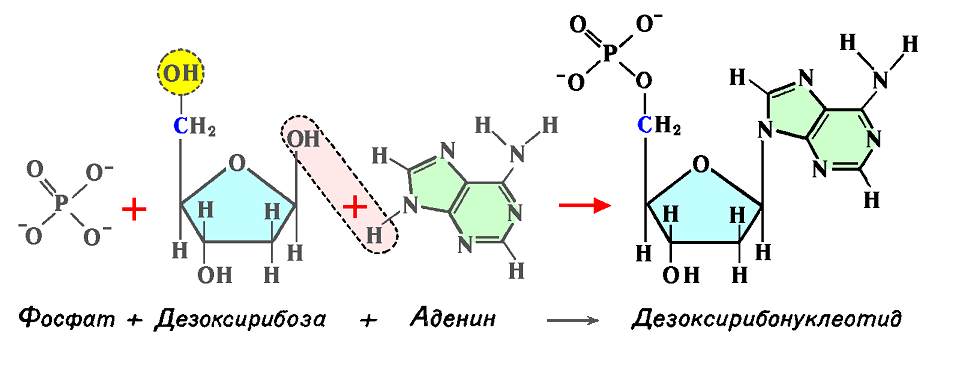
Рис. 5.12. Образование дезоксирибонуклеотида путём соединения фосфата, дезоксирибозы и азотистого основания
Нуклеотиды – это мономеры, из которых строится полинуклеотидная цепь. Соединение друг с другом двух нуклеотидов дает динуклеотиды, трех – тринуклеотиды, затем – тетрануклеотиды, и так вплоть до цепи из сотен тысяч нуклеотидов в виде длинных линейных, неразветвленных полинуклеотидов.
Полинуклеотидные молекулы РНК имеют молекулярную массу 1, 5-2, 0 млн. и состоят из 4-6 тыс. нуклеотидов. Полинуклеотиды ДНК – это обычно гигантские, органические молекулы, имеющие тысячи, миллионы и даже миллиарды нуклеотидов. Последовательность нуклеотидов в цепи молекулы является первичной структурой молекулы ДНК (рис. 5.13).
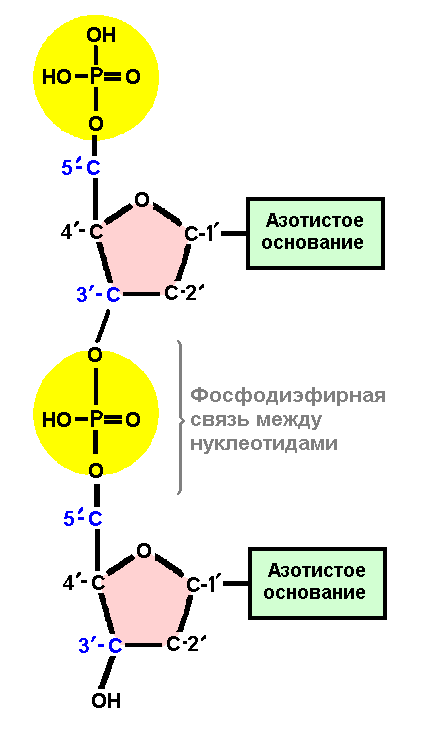
Рис. 5.13. Первичная структура ДНК. Схема соединения нуклеотидов в полинуклеотидную цепь
В молекулах ДНК две полинуклеотидные цепи имеют противоположное направление в отношении связей 5'–3' и 3'–5', т.е. они антипараллельны (рис. 5.14).
Таким образом, в структурной организации молекулы ДНК выделяют три уровня:
– первичную структуру – последовательность нуклеотидов в полинуклеотидной цепи
– вторичную структуру – две комплементарные друг другу и антипараллельные полинуклеотидные цепи, соединенные водородными связями (рис. 5.14);
– третичную структуру – трехмерную спираль с определёнными пространственными характеристиками (рис. 5.11).

Рис. 5.14. Вторичная структура ДНК
Водородные связи между парами комплементарных нуклеотидов (две для пары А-Т и три для пары Г-Ц) относительно непрочные.
Поэтому комплементарные нити молекулы ДНК могут разделяться и соединяться вновь при изменении некоторых условий (например, изменении температуры или концентрации солей).
Разделение двухцепочечной ДНК называется денатурацией, а обратный процесс - образование двухцепочечной структуры ДНК – гибридизацией.
Цепь, содержащая информацию о строении белка (в направлении 5'-3'), называется смысловой цепью, а комплементарная - антисмысловой.
Антисмысловая цепь имеет большое значение при стабилизации структуры двойной спирали ДНК и участвует в процессах репликации и репарации (восстановления) поврежденных участков ДНК.
Молекулы ДНК являются гигантскими полимерами. Единицами измерения длины молекулы приняты: пары нуклеотидов (п.н.). У человека гаплоидный набор содержит 3, 2х109 пар нуклеотидов.
Почти вся ДНК клетки содержится в ядре в виде 46 плотно упакованных, суперскрученных за счет взаимодействий с ядерными белками, структурах - хромосомах. Сравнительно небольшая часть ДНК (около 5%) локализована в митохондриях.