Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Основные проводящие пути ствола мозга
|
|

Рис. 9. Корково-ядерный и корково-спинномозговой пути.
Стрелками показано направление движения нервных импульсов
1 – прецентральная извилина;
2 – таламус;
3 – корково-ядерный путь;
4 – поперечный разрез спинного мозга;
5 – поперечный разрез моста;
6 – поперечный разрез продолговатого мозга;
7 – перекрест пирамид;
8 – латеральный корково-спинномозговой путь;
9 – поперечный разрез спинного мозга;
10 – передний корково-спинномозговой путь.
Восходящие пути:
1. Специфическая восходящая система
· от проприоцепторов в специфические ядра таламуса, а затем в моторную зону коры больших полушарий;
· от слуховых рецепторов – через нижние бугры четверохолмия к медиальному коленчатому телу таламуса, а затем в слуховую зону коры больших полушарий;
· от зрительных рецепторов – через верхние бугры четверохолмия к латеральному коленчатому телу таламуса, а затем в зрительную зону коры больших полушарий.
2. Неспецифическая восходящая система — спиноталамические и спиноретикулярные пути. Импульсы, в основном, от болевых и температурных рецепторов, переключаются в ядрах РФ и неспецифических ядрах таламуса, а затем в различные зоны коры больших полушарий.
3. Пути, по которым импульсы поступают в мозжечок. Через ствол в мозжечок проводится импульсация от проприоцепторов, а также от вестибулярных рецепторов. Затем информация передается в ядра таламуса и далее – в моторную и сенсорную зоны коры больших полушарий.
Нисходящие пути ствола мозга:
§ пирамидный путь – от моторной зоны коры больших полушарий к передним рогам спинного мозга, обеспечивая произвольное сокращение мышц туловища, шеи, головы;
· красноядерно-спинномозговой (рубро-спинальный) путь — от «красного ядра» нервные импульсы направляются в двигательные ядра передних рогов спинного мозга; основная их функция – регуляция мышечного тонуса, равновесия и позы;
· ретикуло-спинальные - от ядер ретикулярной формации ствола мозга к двигательным ядрам передних рогов спинного мозга, участвуют в регуляции тонуса мышц;
· нисходящие пути от коры больших полушарий – к мозжечку, затем обработанная информация поступает в четверохолмие, обеспечивая двигательные реакции организма в ориентировочных реакциях.
Промежуточный мозг
Промежуточный мозг расположен между средним мозгом и полушариями мозга, вокруг Ш-ьего мозгового желудочка. Он состоит из таламической области и гипоталамуса. Таламическая область – главным образованием является – таламус (зрительные бугры); шишковидное тело – эпифиз - орган внутренней секреции, одна из функций – синхронизация биоритмов организма с ритмами внешней среды; коленчатые тела – подкорковые ядра зрительного и слухового анализаторов.
Таламус – является коллектором (собирает) всех афферентных путей (за исключением обонятельных), идущих к большим полушариям. Через таламус проходит вся информация от рецепторов, воспринимающих раздражения из внешней и внутренней среды организма.
Таламус делится прослойками белого вещества на три области – передняя, латеральная и медиальная. Каждая из них представляет собой скопление ряда ядер, различают около 40 ядер таламуса.
Функционально все ядра таламуса делятся на две большие группы – специфические и неспецифические. Специфические ядра таламуса имеют прямые связи с определенными участками коры больших полушарий. Неспецифические ядра в большинстве случаев передают сигналы в подкорковые ядра, от которых импульсы идут одновременно в разные отделы коры больших полушарий.
Специфические ядра таламуса делятся на две группы: переключающие и ассоциативные ядра.
Главными переключающими ядрами являются: 1) латеральное коленчатое тело - переключающее ядро зрительных сигналов, к нейронам этого ядра поступают импульсы от рецепторов сетчатки и верхнего четверохолмия, затем импульсы из таламуса поступают в зрительную зону коры больших полушарий; 2) медиальное коленчатое тело - переключающее ядро слуховых сигналов, импульсы в это ядро поступают от рецепторов улитки внутреннего уха и нижнего четверохолмия, а из таламуса поступают в слуховую зону – коры больших полушарий; 3) заднее вентральное ядро таламуса – к нему приходят импульсы по спинно-таламическому пути (кожная чувствительность), по пучку Голля от рецепторов опорно-двигательного аппарата. Нейроны этого ядра передают информацию в заднюю центральную извилину коры больших полушарий — в сенсорную зону. В заднее вентральное ядро поступает информация также от вкусовых рецепторов; 4) передние ядра таламуса – получают импульсы от обонятельных рецепторов и висцерорецепторов, а затем импульсы передаются в лимбическую область коры больших полушарий.
Ассоциативные ядра таламуса получают импульсы от переключающих ядер таламуса и передают их в ассоциативные зоны коры больших полушарий.
Неспецифические ядра таламуса – некоторые авторы рассматривают эти ядра, как часть ретикулярной формации. Неспецифическая система таламуса принимает участие в быстрой и кратковременной активации коры больших полушарий в противоположность медленной и длительной активации, осуществляемой ретикулярной формацией ствола мозга.
Неспецифические ядра таламуса принимают участие в осуществлении конкретных рефлексов, в частности, они принимают участие в организации процессов внимания у бодрствующего организма, в отличие от ретикулярной формации ствола мозга, которая выполняет функции поддержания тонуса всей коры.
Неспецифические ядра таламуса имеют широкие связи с переключающими и ассоциативными ядрами таламуса, а также с подкорковыми образованиями.
Таламус имеет большое значение в возникновении ощущений, т.к. он передает коре больших полушарий импульсы, возникающие при раздражении различных рецепторов, а также потому, что в ядрах самого таламуса происходит переработка получаемой информации, в результате которой изменяется характер ощущений. Таламус является высшим центром болевой чувствительности. Иногда при поражении таламуса возникает нарушение восприятия болевых ощущений и самые незначительные раздражения (прикосновения) вызывают приступ тяжелейших болей. А иногда наступает состояние – анальгезии - когда болевые раздражения не вызывают ощущений боли.
Гипоталамус находится в основании головного мозга и составляет стенки Ш-его мозгового желудочка. Стенки к основанию переходят в воронку, которая заканчивается гипофизом. Гипоталамус является центральной структурой лимбической системы мозга и выполняет многообразные функции. Часть этих функций относится к гормональной регуляции, которая осуществляется через гипофиз. Другие функции связаны с регуляцией биологических мотиваций (потребление пищи и поддержание массы тела, потребление воды и водно-солевого баланса в организме, регуляция температуры тела в зависимости от температуры внешней среды, функция размножения), эмоциональные переживания, мышечная работа и другие факторы. Гипоталамус играет важную роль в реакции организма на стрессовые воздействия.
Несмотря на то, что гипоталамус занимает не очень большую площадь, он имеет в своем составе 48 ядер. В составе гипоталамуса имеются нейроны, вырабатывающие гормоны или специальные вещества, которые в дальнейшем, действуя на клетки соответствующих эндокринных желез, приводят к выделению или прекращению выделения гормонов. Все эти вещества вырабатываются в нейронах гипоталамуса, затем транспортируются по их аксонам в гипофиз. Ядра гипоталамуса связаны с гипофизом гипоталамо-гипофизарным трактом, который состоит из примерно 200 000 волокон.
Гипоталамус является частью промежуточного мозга и одновременно эндокринным органом. В определенных его участках осуществляется трансформация нервных импульсов в эндокринный процесс. В других областях гипоталамуса образуются рилизинг-факторы. Они играют роль стимуляторов.
Аксоны нейронов гипоталамуса проецируются в гипофиз или во многие участки головного мозга. Таким образом, один и тот же нейрон гипоталамуса может выполнять роль нейрогормона и медиатора или модулятора синаптической передачи.
Гипоталамус, в свою очередь, подвержен влиянию ряда других центров головного мозга. Все эти центры образуют лимбическую систему.
Лимбическая система.
Структурылимбической системы расположены в виде кольца на границе новой коры, отделяющей ее от ствола. В лимбическую систему включают образования древней коры (обонятельная луковица, периамигдалярная и препериформная кора), старой коры (гиппокамп, зубчатая и поясная извилины) и подкорковые ядра (гипоталамус, миндалина, перегородка и передние ядра таламуса).
Афферентные входы в лимбическую систему осуществляются от
различных областей головного мозга, а также через гипоталамус от ретикулярной формации ствола мозга. В лимбическую систему поступают импульсы от обонятельных рецепторов.
Рис. 10. Структуры лимбической системы

Одной из главных структур лимбической системы является гипоталамус. Именно через гипоталамус большинство структур лимбической системы объединены. Имеются многочисленные связи между структурами лимбической системы и гипоталамусом.
Эфферентные пути от лимбической системы направляются через гипоталамус на нижележащие вегетативные и соматические центры ствола мозга и спинного мозга.
Структурной особенностью лимбической системы является наличие хорошо выраженных кольцевых нейронных цепей, объединяющих различные ее структуры. После получения информации о внешней и внутренней среде организма, лимбическая система «запускает» вегетативные, соматические и поведенческие реакции, обеспечивая приспособление организма к внешней среде и сохранение внутренней среды на определенном уровне.
Большие полушария головного мозга
Большие полушария головного мозга состоят из парных частей – правого и левого полушарий и соединяющей срединной части — мозолистого тела. В каждом полушарии головного мозга различают три поверхности – медиальную, выпуклую (латеральную) и нижнюю.
Вещество полушарий расположено вокруг боковых желудочков (I и II мозговые желудочки), они неправильной формы, их стенки образованы веществом полушарий. В каждом боковом желудочке различают центральную часть, передние и задние рога. Боковые желудочки сообщаются с Ш-ьим и IV-ым мозговым желудочками; в полостях желудочков находится церебральная жидкость.
Кора больших полушарий головного мозга является высшим, наиболее поздно развивающимся и особо сложным по своей структуре и функциям отделом центральной нервной системы.
Структурными элементами коры больших полушарий являются нервные клетки - нейроны с отростками и клетки нейроглии.
Кора покрывает всю поверхность больших полушарий; ее толщина варьирует от 1, 5 до 3 мм. Общая поверхность обоих полушарий коры у взрослого человека составляет от 1450 до 1700 см3. В коре полушарий насчитывается от 12 до 18 млрд. нейронов. Нервные клетки расположены в несколько слоев, лежащих друг над другом: I слой – молекулярный, беден нервными клетками, образован в основном сплетением нервных волокон; II слой – наружный зернистый – состоит из мелких клеток; Ш слой – имеет большую толщину, чем предыдущие слои и состоит из крупных пирамидных клеток: IV слой – внутренний зернистый, он состоит из мелких клеток, в некоторых участках коры он иногда отсутствует; V слой – состоит из больших пирамидных клеток Беца, от клеток отходит толстый дендрит, который многократно ветвится в верхних слоях, аксон больших пирамидных клеток спускается в белое вещество и направляется к подкорковым ядрам или к спинному мозгу, образуя основной нисходящий пирамидный тракт.
По функциональному признаку нейроны коры больших полушарий могут быть подразделены на три основные группы. К первой группе относятся клетки, на которых оканчиваются аксоны нейронов специфических афферентных путей. Эти клетки могут быть названы сенсорными, они обеспечивают восприятие афферентных импульсов, проходящих в основном в кору больших полушарий из ядер зрительных бугров (таламуса). Ко второй группе относятся клетки, посылающие импульсы в лежащие ниже отделы мозга, т.е. к подкорковым ядрам, стволу мозга, к спинному мозгу – пирамидные нейроны они в основном сконцентрированы в V слое коры, они еще называются моторными или эфферентными нейронами. Третья группа клеток – это клетки, которые осуществляют связь между различными нейронами, к ним относятся мелкие и средние пирамидные, а также веретенообразные.
В коре больших полушарий имеется также сложная структура нервных волокон. В коре различают горизонтальные волокна, соединяющие различные участки коры и радиальные, связывающие различные участки коры и радиальные, связывающие серое и белое вещества.
По функции волокна больших полушарий (белое вещество) делятся на три группы: ассоциативные – связывают участки коры в пределах одного полушария, комиссуральные - соединяют симметричные участки полушарий через мозолистое тело, проекционные – связывают кору с нижележащими проекционными центрами.
В белом веществе кроме волокон расположены ядра из серого вещества.
Вся поверхность коры изрезана бороздами и между ними расположены извилины. Наиболее крупные борозды: центральная (Роландова борозда), теменно-затылочная, латеральная (височная). Несколько извилин образуют доли полушарий, которые лежат между крупными бороздами – лобная, теменная, затылочная, височная, островок – в глубине боковой извилины.
В кору больших полушарий поступают афферентные импульсы от всех рецепторов организма. Непосредственно передаточной станцией этих импульсов к коре (за исключением импульсов от обонятельных рецепторов) являются ядра таламуса. Участки коры, в которых преимущественно поступают афферентные импульсы – корковые отделы анализаторов — сенсорные зоны коры больших полушарий.
 Рис. 11. Карта архитектонических полей коры головного мозга
Рис. 11. Карта архитектонических полей коры головного мозга
а – первичные поля;
б – вторичные;
в – третичные.
Основная часть информации об окружающей среде и внутренней среде организма поступает в первичные поля сенсорных зон (проекционные поля). Первичные поля, главным образом, состоят из мономодальных нейронов и формируют ощущение одного качества. Затем для дальнейшей обработки информация поступает во вторичные сенсорные поля. В эти области импульсы поступают из проекционных полей и из проекционных (разных) ядер таламуса. Вторичные поля располагаются по краю первичных сенсорных полей. Важной особенностью клеток ассоциативных зон является их полимодальность, т.е. реагируют на раздражение разных рецепторов.
Соматосенсорная зона коры (поле 1, 2, 3) расположена в задней центральной извилине (рис.11). К этой зоне поступают афферентные импульсы от заднего вентрального ядра таламуса. Эти поля получают информацию от кожных (тактильных и температурных) рецепторов и проприоцепторов (суставно-мышечных) противоположной стороны тела.
В соматосенсорной коре имеются зоны проекции различных частей тела человека. Наибольшую площадь занимает корковое представительство рецепторов кисти рук, голосового аппарата и лица, наименьшую площадь – представительство туловища, бедра, голени.
Зрительная зона коры – расположена в затылочной области коры больших полушарий. Афферентные импульсы поступают в эту зону от наружных (латеральных) коленчатых тел таламуса. Здесь имеется представительство рецепторов сетчатки (поле 17). Вокруг этого поля расположены вторичные поля (поля 18, 19), нейроны этих полей полимодальны и они отвечают не только на световые раздражения, но и на тактильные, слуховые, что обуславливает возникновению сложных зрительных образов.
Рис. 12. Схема расположения центров речи:

а – верхнелатеральная поверхность левого полушария; б – медиальная поверхность правого полушария;
1 – ядро кожного анализатора (тактильная, болевая и температурная чувствительность) – в заднецентральной извилине и в верхней теменной дольке;
2 – ядро двигательного анализатора – в предцентральной извилине и околоцентральной дольке;
3 – речедвигательный анализатор – в задней части нижней лобной извилины - «двигательный центр речи» Брока, односторонний, у «правшей» - в левом полушарии, у «левшей» - в правом полушарии;
4 – ядро слухового анализатора - в средней части верхней височной извилины, на поверхности, обращенной к островку – извилины Гешля;
5 – ядро слухового анализатора устной речи – в задней части верхней височной извилины, центр Вернике; 6 – ядро зрительного анализатора, по краям шпорной борозды; 7 – ядро обонятельного и вкусового анализаторов – в крючке.
Слуховая зона коры – расположена в височной доле коры больших полушарий (поле 41, 42, 22). Афферентные импульсы поступают от клеток внутренних (медиальных) коленчатых тел таламуса и несут информацию от слуховых рецепторов улитки внутреннего уха (рис.11). К проекционной коре височной доли относится и корковый отдел вестибулярного анализатора.
В височной доле коры больших полушарий расположен центр Вернике (средняя треть верхней височной извилины) – центр сенсорной речи (поле 42). Повреждения или заболевания сенсорного центра речи приводят к нарушению звукового анализа речи. Возникает сенсорная афазия, при которой становится невозможным различение на слух элементов речи, а, следовательно, понимание речи, хотя способность различать неречевые звуки остаются нормальными.
На границе височной, затылочной и теменной расположен центр чтения и распознавания и хранения образов письменной речи - (поле 39), который тесно связан с центром Вернике, с центром зрительного анализатора в затылочной области и с центрами в теменной области.
А в заднем отделе второй и третьей лобных извилин расположен моторный (двигательный) центр речи – центр Брока (поле 44). Повреждения и заболевания моторного центра речи ведут к нарушению анализа и синтеза кинестетических (двигательных) раздражений, возникающих при произнесении звуков речи. Наступает моторная афазия, при которой становится невозможным произнесение слов, фраз, хотя движения речевых органов (движение языка и губ, открывание и закрывание рта и т.д.) не нарушаются. Больные неправильно произносят слова, меняют в них последовательность звуков.
Представительство вкусовой рецепции (в самой нижней части саматосенсорной зоны коры в области представительства языка, в крючке и аммоновом роге – поле 43) – афферентные импульсы поступают от нижнего заднего ядра таламуса. Обеспечивает проекционную функцию и распознавание вкусовых образов.
Представительство обонятельного анализатора (передняя часть грушевидной доли коры в области извилины морского конька – поле 11) – импульсы от обонятельных рецепторов полости носа идут по обонятельному тракту до передней части грушевидной доли. Обеспечивает хранение и распознавание обонятельных образов.
Моторные зоны коры больших полушарий – находятся спереди от центральной извилины (рис.11). В первичной моторной коре (поле 4) расположены нейроны, иннервирующие мышцы лица, туловища и конечностей. В ней имеется четкая проекция мышц тела. Раздражение первичной моторной коры вызывает сокращение мышц противоположной стороны тела. При поражении этой корковой зоны утрачивается способность к тонким координированным движениям конечностями и особенно пальцами рук. Пространственная организация моторной зоны у человека неравномерная. Выше всех расположено представительство нижних конечностей, еще ниже – двигательные точки туловища, еще ниже – верхние конечности, еще ниже – мускулатура головы. В моторной зоне, как и в сенсорной, наибольшую площадь занимает мускулатура кистей рук, лица, губ, языка и наименьшую – туловища и нижних конечностей.
Нарушения в моторной области коры несут за собой у человека полный или частичный паралич мускулатуры противоположной половины тела. Наиболее сильно страдает при поражении моторной зоны у человека способность к отдельным мелким движениям, например, пальцев рук или ног.
Рис. 13. Схема чувствительного и двигательного гомункулусов
( Разрез полушарий во фронтальной плоскости)
 А – проекция общей чувствительности в коре постцентральной извилины
А – проекция общей чувствительности в коре постцентральной извилины
1 – голова; 2 – рука; 3 – кисть;
4 – пальцы; 5 – глаз; 6 – лицо;
7 – губы; 8 – челюсть и зубы;
9 – язык; 10 – глотка;
11- половые органы;
12 – стопа; 13 – нога; 14 – таз;
15 – торс; 16 - шея
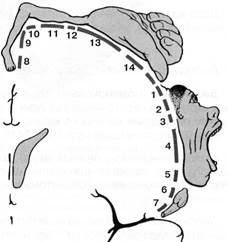 Б – проекция двигательной системы в коре предцентральной извилины
Б – проекция двигательной системы в коре предцентральной извилины
1 – лоб; 2 – глаз и веко; 3 – лицо;
4 – мимические мышцы; 5 – челюсть;
6 – язык; 7 – глотка;
8 – стопа; 9 – нога; 10 – таз;
11 – торс; 12 – рука;
13 – кисть; 14 – пальцы.
Часть отростков пирамидных клеток моторной коры образуют кортикоспинальный тракт (пирамидный). Другая часть нисходящих нервных волокон направляется из моторной зоны в подкорковые структуры, образуя два вида путей. Одни из них связывают клетки моторной зоны с полосатым телом (подкорковые ядра), красным ядром и черной субстанцией (средний мозг). Другие – идут через мост, обеспечивая связь моторной коры с мозжечком. Часть нисходящих волокон моторной зоны коры направляется непосредственно к клеткам ретикулярной формации и гипоталамуса, что обуславливает сосудистые реакции в ответ на раздражение моторной зоны коры.
Посылая импульсы по всем этим путям к лежащим ниже отделам ЦНС, моторная зона коры больших полушарий управляет деятельностью двигательного аппарата организма.
Вторичная моторная кора (поле 6) имеет главенствующее функциональное значение по отношению к первичной двигательной коре, осуществляя высшие двигательные функции, связанные с планированием и координацией произвольных движений. Раздражение коры поля 6 (рис.11) вызывает сложные координированные движения, например поворот головы, глаз, туловища в противоположную сторону, содружественное сокращение сгибателей и разгибателей. Здесь также расположен центр автоматизма письма, который тесно связан с центром Брока.
Ассоциативные области коры – включают участки новой коры больших полушарий, которые расположены вокруг проекционных зон и рядом с двигательными зонами, но они не выполняют непосредственно чувствительных или двигательных функций. У человека они составляют около 50% всей коры. К ассоциативной коре относят теменно-височно-затылочную кору, префронтальную кору лобных долей, лимбические ассоциативные зоны. Нейроны ассоциативной коры полисенсорны, они отвечают на несколько раздражителей. Эти зоны связаны с различными проекционными полями и с ассоциативными ядрами таламуса.
В настоящее время выделяют две основные ассоциативные системы: таламотеменную и таламолобную.
Таламотеменная система — основными функциями являются гнозис (узнавание) формы, величины, значение предмета, понимание речи и праксис – целенаправленное действие, хранение и реализация программ двигательных автоматизированных актов.
Таламолобная система - основная роль сводится к формированию функциональных систем целенаправленных поведенческих актов. Эта система играет важную роль в выработке стратегий поведения. Нарушение этой функции особенно заметно, когда необходимо быстро изменить действие и когда накопилась серия раздражителей и требуется выбрать правильное решение для поведенческой реакции.
Задание для самоподготовки
1) Каково структурирование нервной системы по пространственному и функциональному признакам?
2) Назовите отделы головного мозга, где проходит граница между ними.
3) Перечислите ядра черепно-мозговых нервов, расположенных в толще ствола мозга.
4) Где расположены центры жизненно важных безусловных рефлексов.
5) Назовите основные двигательные ядра ствола мозга, и с какими выше- и нижележащими структурами головного мозга они связаны.
6) Перечислите симптомы расстройств двигательных функций при мозжечковых нарушениях.
7) Назовите основные восходящие и нисходящие пути ствола мозга.
8) Дайте характеристику понятию “цитоархитектоника“ коры головного мозга.