Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Вегетативные и покоящиеся формы микроорганизмов, морфо-функциональная характеристика. Виды покоящихся форм микробов. Условия образования покоящихся форм.
|
|
образованию эндоспор способна только небольшая группа бактерий. Огромное значение эндоспор связано с их термоустойчивостью. В то время как почти все остальные бактерии, а также вегетативные клетки спорообразующих видов гибнут при 80°С (при температуре пастеризации) через 10 мин, терморезистентные эндоспоры переносят значительно более сильный нагрев; некоторые споры выдерживают даже кипячение на протяжении многих часов. Трудоемкая и дорогостоящая техника стерилизации (обеззараживания) рассчитана на уничтожение эндоспор. С другой стороны, терморезистентность спор предоставляет своеобразную возможность избирательного обогащения культур спорообразующих форм. Землю или иной материал, взятый в определенном месте, на протяжении 10 мин прогревают при 80 или 100°С, что приводит к гибели вегетативных клеток; только терморезистентные споры остаются жизнеспособными и прорастают в подходящей питательной среде.
Классификация бактерий, образующих эндоспоры. Спорообразующие виды, за одним исключением, относятся к палочковидным грам-положительным бактериям. Большинство из них подвижно благодаря перитрихально расположенным жгутикам. Бактерии, принадлежащие к роду Bacillus, - строгие аэробы или факультативные анаэробы. Роды Clostridium и Desulfotomaculum объединяют анаэробные бактерии, способные к образованию спор. Клостридии получают энергию за счет брожения; виды Desulfotomaculum способны получать энергию путем анаэробного дыхания, используя сульфат в качестве акцептора электронов. Sporolactobacillus принадлежит к числу молочнокислых бактерий. Sporosarcina имеет сферические клетки, но по своим физиологическим признакам относится к бациллам. Содержание GC в ДНК клеток, способных образовать споры, удивительно низко; клостридии, у которых ДНК содержит от 22 до 27 мол. % GC, оказываются в конце списка прокариот, расположенных в порядке убывания этого показателя.
Выявление эндоспор. При микроскопическом исследовании споры видны благодаря своему высокому показателю преломления - такому же, как у обезвоженного белка; это указывает на то, что в спорах большое количество богатого белком материала сконцентрировано в малом объеме. Спора содержит почти все сухое вещество материнской клетки, но занимает в 10 раз меньший объем. В сомнительных случаях вопрос о том, имеются ли в клетках истинные эндоспоры, можно решить с помощью специального окрашивания. Если препарат бактерий, фиксированный нагреванием, прокипятить с карболовым раствором фуксина, то споры прочно связывают краситель и не обесцвечиваются даже при обработке этанолом или 1 М уксусной кислотой, тогда как остальное содержимое клетки при этом становится бесцветным.
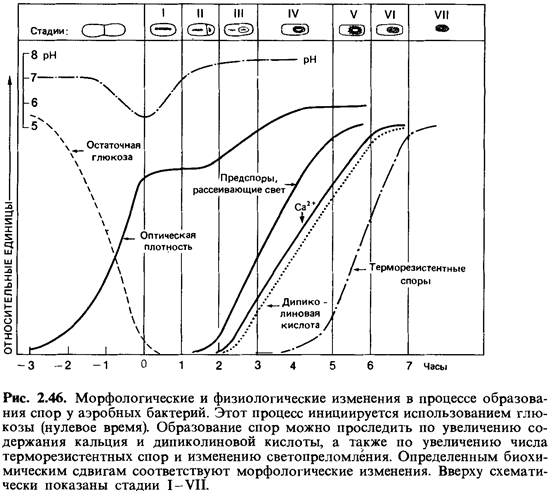
Спорообразование (споруляция). Споры образуются внутри бактериальной клетки. Этот процесс начинается с накопления белкового материала, поэтому показатель преломления в месте образования споры возрастает. Происходящие при этом метаболические превращения сопровождаются расходованием запасных веществ (поли-3-гидроксимасляной кислоты у аэробов и полисахаридов у анаэробов). В течение первых пяти часов спорообразования значительная часть белков материнской клетки распадается. При этом образуется специфичное для спор вещество - дипиколиновая (пиридин-2, 6-дикарбоновая) кислота. В вегетативных клетках эта кислота не встречается. В ходе синтеза дипиколиновой кислоты происходит поглощение ионов кальция; в зрелых спорах эта кислота находится, по-видимому, в виде хелата с кальцием и может составлять 10-15% сухого вещества спор. Дипиколиновая кислота локализована в протопласте споры и имеется только в терморезистентных эндоспорах (рис. 2.45, 2.46).
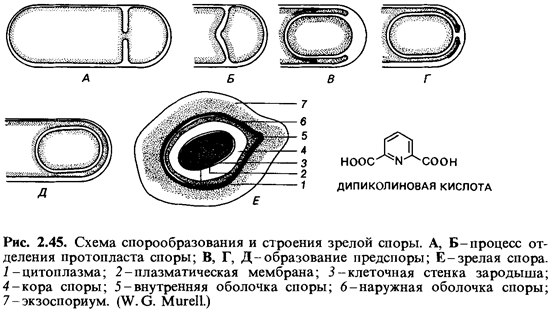
Спорообразование - один из сложнейших процессов дифференцировки бактериальной клетки. Оно начинается с особого неравного деления клетки (рис. 2.45). В результате впячивания плазматической мембраны часть протопласта отшнуровывается от материнской клетки. Этот протопласт содержит часть ядерного материала - один геном. Образования клеточной стенки между обоими протопластами (как при обычном делении) здесь не происходит. Вместо этого протопласт будущей споры окружается, как бы обрастает, плазматической мембраной материнской клетки. В результате вокруг него оказываются две плазматические мембраны, и каждая из них участвует в синтезе стенки споры. Мембрана протопласта споры синтезирует снаружи от себя стенку зародышевой клетки, а мембрана, происходящая от материнской клетки, синтезирует вовнутрь кору споры (кортекс). Последняя состоит из многослойного пептидогликанового остова, который отличается от каркаса стенок вегетативных клеток помимо прочего степенью сшивки. Наружную оболочку споры образует материнская клетка; эта оболочка в значительной части состоит из полипептидов. Материнская же клетка образует еще один дополнительный тонкий полипептидный покров - экзоспорий; он имеется только у немногих бактерий (например, у Bacillus cereus) и окружает спору в виде свободного чехла. Ввиду такой многослойности покровов не покажется удивительным, что на долю оболочки приходится около половины объема и соответственно сухой массы зрелой споры.
Индукция спорообразования. Споры отнюдь не являются обязательной стадией жизненного цикла бацилл; при благоприятных условиях питания бациллы могут неограниченное время размножаться делением как вегетативные клетки. Образование спор начинается лишь тогда, когда не хватает питательных веществ или когда в избытке накапливаются продукты обмена. Иными словами, оно происходит лишь в тех случаях, когда этому способствуют общие условия. Высыхание не стимулирует споруляцию. Если поместить вегетативные клетки в дистиллированную воду, можно наблюдать «эндотрофную споруляцию», т.е. образование спор за счет накопленных внутри клеток запасных веществ. В таких случаях образование спор, очевидно, вызывается недостатком экзогенного субстрата. Индукция спорообразования осуществляется в течение не скольких часов. Если, например, к суспензии вегетативных клеток Bacillus cereus var. mycoides в первые 5 ч после помещения клеток в воду добавить глюкозу, то образование спор прекратится - добавление субстрата подавляет споруляцию. Глюкоза, добавленная позднее чем через 6 ч, менее действенна: подавление спорообразования не происходит. Индукция (дерепрессия) спорообразования продолжается, и через 10-13 ч после переноса клеток в воду около 90% их образует споры. Следовательно, споруляция регулируется внешними факторами.
Число клеток, образующих споры, во многих случаях возрастает после добавления к среде солей марганца.
Способность образовывать эндоспоры постепенно утрачивается при многократных пересевах вегетативных клеток. Поскольку суспензии спорообразующих микроорганизмов обычно содержат и споры, и вегетативные клетки, перед каждым пересевом культуру, как правило, подвергают кратковременному кипячению. Это способствует сохранению или повышению способности клеток образовывать споры.
Свойства зрелых спор. Споры освобождаются при автолизе материнских клеток. Зрелые споры не проявляют никакой метаболической активности. Они чрезвычайно устойчивы к воздействию высокой температуры, разного рода излучений и химических агентов. Терморезистентность обусловлена очень низким содержанием воды. В спорах Bacillus megaterium всего лишь около 15% воды, т.е. примерно столько же, сколько в шерсти или сухом казеине. Лиофилизированные вегетативные клетки бактерий тоже очень термостойки. Терморезистентность спор приблизительно пропорциональна содержанию в них дипиколиновой кислоты.
Радиорезистентность спор также выше, чем у вегетативных клеток. Она примерно пропорциональна содержанию дисульфидных групп в наружном слое белка. Оболочка споры содержит главным образом белок, богатый цистеином и напоминающий кератин. Химическая же устойчивость эндоспор обусловлена непроницаемостью их оболочки для многих веществ.
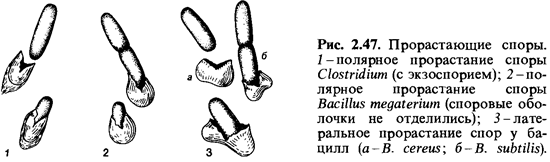
Прорастание спор. Вподходящих средах большинство спор прорастает. Соответствующая предварительная обработка, определенные условия хранения и прогрев могут повысить «всхожесть» спор - увеличить процент прорастания. В случае Bacillus subtilis оптимальными условия ми для стимуляции прорастания спор считают семидневный период покоя и пятиминутный прогрев в воде при 60°С. Другие споры могут быть активированы кратковременным кипячением (10 мин при 100°С). Обработка тепловым шоком должна проводиться непосредственно перед высевом спор, так как процесс активации обратим. Прорастанию спор предшествует поглощение ими воды и набухание. Для прорастания активированных спор в ряде случаев необходимо присутствие глюкозы, аминокислот, нуклеозидов или других соединений. В ходе прорастания спор происходят глубокие физиологические изменения: дыхание и ферментативная активность быстро возрастают; начинается выделение аминокислот, дипиколиновой кислоты и пептидов. При прорастании спор потеря сухого вещества достигает 25-30%. Во время прорастания споры теряют свою термоустойчивость. Выходящая из споры ростовая трубка бывает окружена очень тонкой и, видимо, неполностью сформированной клеточной стенкой, так что в протопласт может, например, проникать даже ДНК. Ростовая трубка может образоваться как в полярном, так и латеральном положении; в одних случаях оболочка споры при этом разрывается, в других ростовая трубка прокалывает ее (рис. 2.47).
Продолжительность жизни спор. Бактерии в виде спор могут дли тельное время находиться в состоянии анабиоза. В земле, которая прилипла к растениям из гербария Кью Гарденс (Англия) и пролежала вместе с ними в сухом состоянии от 200 до 320 лет, было обнаружено лишь небольшое количество жизнеспособных спор Bacillus subtilis и В. liche-niformis. В пробах почвы, хранившихся от 50 до 100 лет, были также обнаружены споры В. coagulans и В. circulans. По данным такого рода экспериментов, в сухой почве за 50 лет хранения до 90% спор теряет жизнеспособность. Судя по этому, одна тонна сухой почвы и через 1000 лет все еще будет содержать жизнеспособные споры.
В сухом состоянии многие бактерии (если не большинство из них) на протяжении ряда лет сохраняют жизнеспособность. Для консервации бактерий в коллекциях вегетативные клетки подвергают, как правило, лиофильной сушке и хранят при комнатной температуре или при низких температурах в вакууме. Как рассчитал еще Беккерель, микроорганизмы при температуре, близкой к абсолютному нулю, могут оставаться жизнеспособными на протяжении миллионов лет. Кратковременные опыты с жидким азотом и экстраполяция их результатов позволяют заключить, что такие предположения вполне оправданны. Бактерии, не переносящие лиофилизации, выдерживают многолетнее хранение в суспензиях при температуре жидкого азота.
Другие покоящиеся формы (цисты, экзоспоры, миксоспоры). Эндоспоры являются длительно переживающими формами бактерий, устойчивыми к высокой температуре, высыханию, облучению и химическим воздействиям. Помимо эндоспор у некоторых других бактерий существуют иные покоящиеся формы - экзоспоры и цисты. Образование экзоспор до сих пор наблюдалось только у метаболизирующих метан бактерий Methylosinus trichosporium. Экзоспоры возникают путем почкования материнской клетки; они сходны по своим свойствам с эндоспорами бацилл. Некоторые бактерии образуют шарообразные толстостенные клетки, называемые цистами. При истощении пищевых ресурсов в цисту превращается вся палочковидная вегетативная клетка, а не только часть ее, как при образовании эндоспоры. Цисты видов Azotobacter, а также Methylocystis устойчивы к высушиванию, механическим нагрузкам и облучению, но не к действию высоких температур. Подобного рода превращение всей клетки в цисту лежит и в основе образования миксоспор из палочковидных вегетативных клеток Myxococcus и Sporocytophaga.
Клетки Arthrobacter (A. globiformis) плеоморфны. При обилии субстрата они растут в виде палочек; когда же субстрат иссякает, появляются кокковидные клетки. Arthrobacter относится к тем бактериям, которые способны некоторое время сохраняться в высохшей почве, пре бывая в состоянии покоя; структурной дифференцировки в данном случае не выявлено.