Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Распознавание грибов
|
|
К сожалению, в настоящее время грибы остаются недостаточно исследованной группой патогенов в плане распознавания их элементов системой врожденного иммунитета. Однако основные соединения – мишени в структуре клетки грибов приведены на рисунке 14 и таблице 7.

Рисунок 14. Структура клеточной стенки грибов.
Рецепторы системы врожденного иммунитета, распознающие грибы, включают эффекторные гуморальные факторы: ферменты (хитиназа), опсонины (С3, пентраксины), клеточные рецепторы для фагоцитоза (МР, SP-A, SP-D), сигнальные клеточные рецепторы (ТПР4, 2, 9, МР).
Таблица 7. Распознавание грибов рецепторами системы врожденного иммунитета
| Рецептор/эффекторная молекула | Лиганд |
| Хитиназа | Хитин |
| Хитотриозидаза | Хитин |
| CD14 | Глюкуроноксиманнан |
| С3 | Поверхность клетки |
| С3R | Манноза, бета-глюкан, N-ацетилглюкозамин, метилманнозид, опсонизированные комплементом грибы |
| DC-SIGN | Манноза |
| Дектин-1 | Бета-глюканы |
| Лактозилцерамид | Бета-глюканы |
| Маннозный рецептор | Терминальная манноза |
| Манноза-связывающий лектин | Моносахариды (манноза, фукоза, глюкоза) |
| Пентраксин 3 | Галактоманнан, зимозан |
| SP-A | Моносахариды (манноза, фукоза, глюкоза) |
| SP-D | Моносахариды (манноза, фукоза, бета-глюкан) |
| ТПР2 | Фосфолипоманнан, зимозан, липопротеины, гликолипиды |
| ТПР4 | Маннан, глюкуроноксиманнан |
| ТПР9 | ДНК |
Подобно другим патогенам, клетки грибов распознаются различными рецепторными молекулами, что ведет к повышению вероятности распознавания (например, криптококки экранинуют бета-глюканы массивной капсулой из глюкуроноксиманнана. Таким образом они защищаются от распознавания через дектин-1 и многих естественных опсонинов, однако материал капсулы распознается ТПР4 и возбуждает сильный воспалительный ответ). Общий вектор активности иммунной системы в ответ на грибковую инфекцию – развитие иммунного воспаления (Th1-зависимого) с мобилизацией ресурсов системы врожденного иммунитета (фагоциты и цитотоксические клетки). Тем не менее, современные исследования подтверждают важную роль антител в патогенезе микозов. Основные позитивные функции антител сводятся к опсонизации, активации комплемента, «фокусировании» механизмов естественного иммунитета. К сожалению, грибы зачастую устойчивы к фагоцитозу и действию комплемента, а некоторые (H.capsulatum) используют указанные механизмы защиты для проникновения в чувствительные клетки.
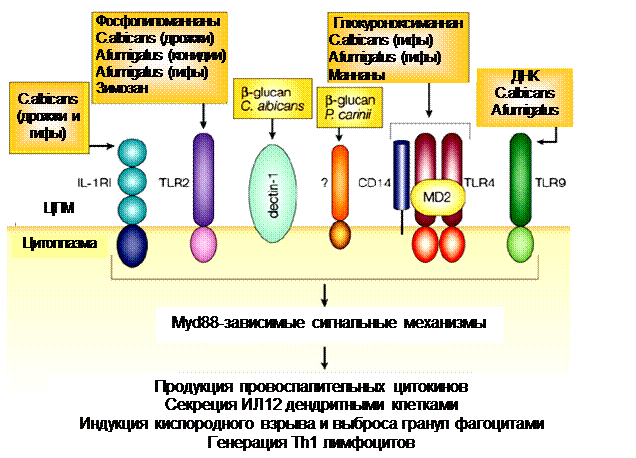
Сложность структуры патогена и множественность распознавания создают возможность для регулирования и «индивидуализации» реагирования со стороны иммунной системы. Так, С.albicans преимущественно распознается тремя рецепторами: ТПР4 связывает О-маннаны, МР – N-маннаны, Дектин-1 - бета-глюканы. При этом ТПР4 возбуждает секрецию хемокинов, мобилизацию нейтрофилов и организацию иммунного воспаления (Th1), что связывают с развитием иммунитета, уничтожением возбудителя и выздоровлением; ТПР2 вызывает «иммунологическое отклонение» - смещение равновесия Th1/Th2 в сторону Th2 (стимуляция гуморальных реакций/аллергии ГНТ) такое развитие инфекционного процесса грозит формированием пермиссивного фенотипа и хронизацией заболевания. В то же время, распознавание через лектиновые рецепторы и рецепторы комплемента играет важную роль в фагоцитозе возбудителя и регулировании воспаления.
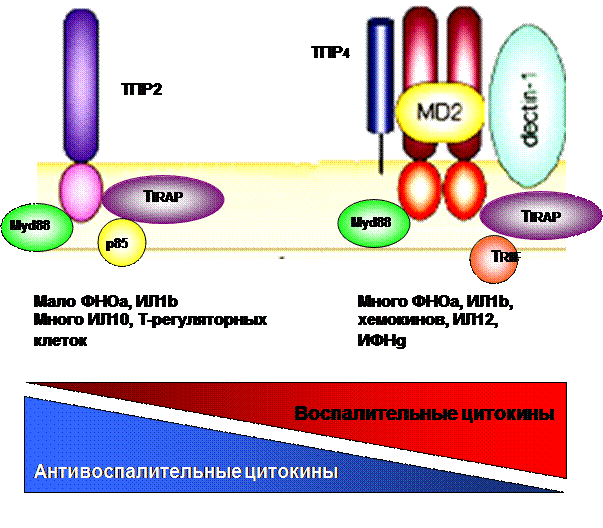
Рисунок 15. Регулирование ответа на C. Albicans посредством экспрессии/распознавания рецепторами врожденного иммунитета
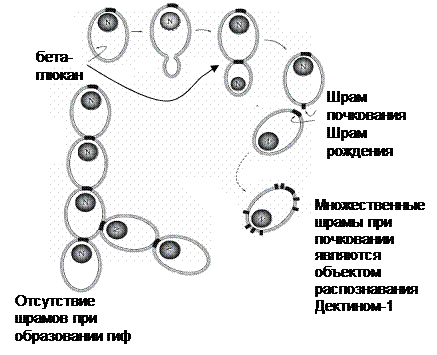 Патогенные грибы, со своей стороны, используют регуляторные механизмы иммунной системы для уклонения от иммунного ответа: например, Aspergillus fumigatus в мицелиальной форме не распознается ТПР4, в то аремя как механизм иммунного отклонения, опосредуемый ИЛ10, продолжает работать и способствует формированию благоприятных условий для развития микоза. Схожий механизм использует С.albicans, для которого переход в мицелиальную форму четко ассоциируется с патогенностью. Кроме уклонения от распознавания ТПР4, гифы кандид не связывают Дектин-1 (бета-глюканы на гифах экранированы слоем маннанов).
Патогенные грибы, со своей стороны, используют регуляторные механизмы иммунной системы для уклонения от иммунного ответа: например, Aspergillus fumigatus в мицелиальной форме не распознается ТПР4, в то аремя как механизм иммунного отклонения, опосредуемый ИЛ10, продолжает работать и способствует формированию благоприятных условий для развития микоза. Схожий механизм использует С.albicans, для которого переход в мицелиальную форму четко ассоциируется с патогенностью. Кроме уклонения от распознавания ТПР4, гифы кандид не связывают Дектин-1 (бета-глюканы на гифах экранированы слоем маннанов).
Другие грибы, напротив, патогенны в дрожжеподобном состоянии: С.neoformans экранирует свои структуры крупной капсулой, а H.capsulatum при переходе в гифальную форму перестраивает бета-глюканы в альфа-глюканы.