Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Структура растительности
|
|
Различают вертикальную и горизонтальную структуру растительного покрова.
Вертикальная структура выражена в форме ярусности. Ярусность проявляется в том, что основная масса листвы у растений разных сосуществующих видов располагается на разной высоте. Этой хорошо заметной надземной ярусности крон и побеговых систем соответствует скрытая в земле ярусность корневых систем. При этом наиболее высокие растения, как правило, имеют и наиболее глубоко проникающие корни, а чем ниже растение, тем более поверхностна его корневая система. Ярусность представляет собой один из способов дифференциации экологических ниш разных видов растений.
Горизонтальная структура растительности (рисунок 9) проявляется в том, что на разных участках преобладают растения разных видов и/или отсутствуют растения каких-либо видов, встречающиеся по соседству. Причины горизонтальной неоднородности растительного покрова бывают разными. В том случае, если она вызвана исходными различиями абиотических факторов среды (влажность, химический состав субстрата, термический режим и т.п.), говорят о комплексности растительности. Неоднородность растительности, обусловленную отмиранием крупных растений в силу разных причин, случайным разносом диаспор, вегетативной подвижностью/неподвижностью растений и т.п., называют мозаичностью. Масштаб мозаичности варьирует в широких пределах.
Несмотря на горизонтальную и вертикальную неоднородность растительности, в ней обычно различимы отдельные участки, имеющие специфический облик. Они кажутся довольно однородными на фоне других подобного рода участков. Такие выглядящие однородными участки растительности существенно различаются по своей природе.
В некоторых случаях однородный участок растительности представляет собой заросль растений одного вида высших растений. Такие одновидовые заросли образует виоленты, которые в определенных условиях оказываются настолько мощными конкурентами, что полностью подавляют другие высшие растения, перехватывая все доступные ресурсы. Примерами могут служить заросли тростника (Fragmites australis) в плавнях или ели обыкновенной (Picea abies) в лесной зоне (так называемые мертвопокровные ельники). Одновидовые заросли могут образовывать и патиенты, например, ризофора (Rhizophora mangle) в тропической мангрове или виды солероса (Salicornia) на литоралях северных морей и по берегам внутриконтинентальных соленых водоемов. Причина появления таких зарослей состоит в том, что на соответствующей территории нет других видов, приспособленных к экстремальному значению определенного фактора внешней среды. Возможно встретить и одновидовые заросли эксплерентов. Так, на песчаных субстратах насыпей дорог, пустырей развиваются заросли щирицы белой {Amamnthus albus), мать-и-мачехи (Tussilago farfara) или якорцев (Tribulus terrestris). Одновидовые заросли эксплерентов появляются потому, что на такие участки случайно попадают диаспоры только одного вида. В отличие от одновидовых зарослей виолентов и патиентов заросли эксплерентов недолговечны. Общую черту любых зарослей составляет слабая пространственная структурированность. Хотя отдельные особи или надземные побеги длиннокорневищных растений располагаются неравномерно, выявить четкую мозаичность не удается. Заросли виолентов одновозрастны, причем размерные различия растений в них невелики и все они располагаются в одном ярусе. Заросли патиентов бывают как одновозрастными (Salicornia), так и разновозрастными (Rhizophora), особи в них заметно различаются высотой, но не составляют четких ярусов.
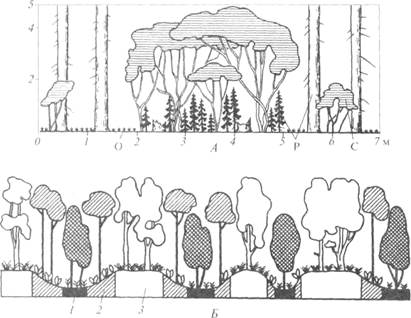
Рисунок 9 – Горизонтальная структура растительности:
А — мозаичность орешниково-елового диаспорического субклимакса; Б — комплексность дубово-ольхово-осинового леса; С — Corylus avellana; О — Oxalis acetosella; P — Picea abies; 1 — ассоциации Alnus incana — Stellaria holostea, 2 — Populus tremula— Convallaria majalis, 3 — Quercus
robur— Carex pilosa
Заросли эксплерентов практически одновозрастны, особи в них часто очень сильно различаются размерами в зависимости от обеспеченности ресурсами, но различить ярусы не удается и в этих зарослях.
Значительно более распространены относительно однородные участки растительности из особей нескольких видов. Они формируются в двух случаях.
Растения видов, у которых преобладают элементы S - и/или R -стратегий, слабо преобразуют среду. При совместном произрастании растений таких видов возникают растительные группировки. Между растениями группировки может возникнуть острая конкуренция, но она не приведет к дифференциации экологических ниш. Растения в группировке приспособлены не столько к сосуществованию друг с другом, сколько к особенностям биотопа. В группировке обычно какой-либо вид преобладает над другими по численности и/или биомассе особей. Такой вид называют доминантом. Какой вид и в какой степени будет доминировать в конкретном местообитании, зависит в основном от случайных причин и сильно варьирует от участка к участку группировки. Мозаичность группировок также случайным образом варьирует в широких пределах. Во многих группировках растения закономерно различаются по высоте побегов и глубине корневых систем, но ярусы часто размытые или вообще незаметны.
Растительное сообщество, или фитоценоз, отличается от группировки тем, что состоит из растений видов, коадаптированных к совместному существованию благодаря дифференциации своих экологических ниш. Среди таких видов особое значение имеет эдификатор — вид, в наибольшей степени преобразующий среду и тем самым определяющий условия жизни растений других видов сообщества. Степень коадаптации видов в сообществе различается. Эдификатор и наиболее тесно коадаптированные с ним виды — ценофилы — составляют основу сообщества и в ненарушенных сообществах всегда присутствуют в полном наборе. В сообществе доминируют ценофильные виды. Наряду с ними встречаются — главным образом в пограничной зоне между сообществами и в местах слабых нарушений сообщества — менее тесно связанные с ними сопутствующие виды, проявляющие определенную степень ценофобности. Их состав может варьировать в разных участках, занятых сообществом. Наименее ценофобные виды, закономерно встречаясь в сообществе, отражают специфику внешней среды сообщества (виды-индикаторы). Более ценофобные виды обитают в группировках и лишь случайно проникают в сообщества в связи с их нарушениями. Чем обильнее в сообществе растения ценофобных видов, тем сильнее нарушено это сообщество.
Растительным сообществам свойственна мозаичность, отражающая особенности жизненного цикла и возобновления доминантов, в первую очередь эдификатора. В большинстве сообществ хорошо заметны ярусы, причем в каждом ярусе имеется собственный доминант. Эдификатор, как правило, доминирует в верхнем ярусе.
Наряду с этими — типичными — растительными сообществами существуют и такие, в которых средообразующую роль играют животные. Например, своеобразными «эдификаторами» степей и прерий являются крупные зеленоядные грызуны, копытные и прямокрылые, а «эдификаторами» некоторых типов саванн — копытные и хоботные.