Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
III Водорастворимые 1 страница
|
|
| Рнбофлави Пантотена! |
Рибофлавин Пантотеньвая
Тиамина дифос-
трн фосфат ФМН, ФМН. Н2; ФАД, ФАД • Hj Пантетенн-4- фосфат, КоА, де- фосфо-КоА
Антиневрнтный Витамин роста
Продолжение табл. 30
|
ний. Эти варианты одного и того же витамина называют витамерами. Они обладают сходным специфическим, но отличающимся1 по силе биологическим эффектом на организм-
Пути метаболизма витаминов в организме. Некоторые витамины поступают с пищей в виде предшественников — провитаминов, которые в тканях превращаются в биологически активные формы витаминов.
Поступающие при всасывании жирорастворимые витамины депонируются в тканях; водорастворимые витамины превращаются в коферменты и, соединяясь с апоферментом, входят в состав сложного фермента. Поскольку срок жизни ферментов ограничен, то коферменты распадаются и выводятся в виде различных метаболитов из организма. Жирорастворимые^ витамины тоже под-
растворимые. Поэтому необходимо постоянное поступление витаминов с пищей.
Нарушение баланса витаминов в организме. Дисбаланс витаминов проявляется в форме недостатка (отрицательный баланс) и избытка (положительный баланс). Частичный недостаток витамина (клинически или биохимически проявляющийся отрицательный баланс) называется гиповитаминозом, а крайне выраженный дефицит — авитаминозом. Недостаток одного витамина относят к моногиповитаминозам, а сразу нескольких — к полигиповитамино- зам. Избыточное накопление в тканях витаминов (выраженный положительный баланс), сопровождающийся клиническими и биохимическими признаками нарушений, называется гипервитаминозом. Он характерен для жирорастворимых витаминов.
Все гиповитаминозы и авитаминозы проявляются задержкой роста молодого организма. Кроме того, для конкретного гиповитаминоза " характерны свои симптомы нарушений обмена веществ и функций, отражающих регулятор ные свойства данного витамина. По этим симптомам выявляют недостаточность соответствующего витамина. Причины гиповитаминозов могут быть экзогенные и эндогенные. К экзогенным относятся- нерациональное питание (однообразная, бедная витаминами пища), изменение состава нормальной кишечной флоры (дисбактериоз), обычно вызываемое длительным применением химиотерапевтических средств (антибиотиков, сульфаниламидов ит. д.); к эндогеиным — нарушения всасывания и транспорта витаминов, образования коферментов (вследствие генетически обусловленных дефектов апофермента или ферментов синтеза коферментов), усиление распада витаминов, физиологически обусловленная высокая потребность в витаминах (растущий организм, беременность).
Гипервитаминоз, или витаминная интоксикация, проявляется общими симптомами: потеря аппетита, расстройство моторной функции желудочно- кишечного тракта, сильные головные боли, повышенная возбудимость нервной системы, выпадение волос, шелушение кожи и некоторые специфические признаки, свойственные данному витамину. Гипервитаминоз может закончиться смертельным исходом.
Причиной гипервитаминозов служит избыточный прием продуктов, богатых данным жирпрястиппнмым витамином (например, печени белого медведя или кита, богатых витамином А), или назначение чрезмерных доз витаминов.
Практическое применение витаминов. В настоящее время применяются отдельные витамины, комбинированные поливитаминные препараты и кофер- ментные препараты витаминов. Последние особенно перспективны, поскольку, будучи введенными в организм, могут сразу участвовать в регуляции обмена веществ. Витамины используются как средство заместительного лечения (при гипо- и авитаминозах) и патогенетического лечения (когда используют отдельные ценные качества витаминов — сосудорасширяющее, регенеративное и т. д.).
2. Жирорастворимые витамины
Ретинол (витамин А)
ного происхождений. Наиболее'богата нм печена различных рыб, особенно трески и морского окуня. Много витамина А в свиной' и говяжьей печени, желтке яиц, сметане, цельном молоке. В растительных продуктах: моркови томатах, свекле, салате содержатся каротиноиды, являющиеся провитамина ми А. Поэтому частично обеспечение витамином А происходит за счет расти тельных продуктов, если в организме не нарушен процесс превращения пищевых каротиноидов в витамин А. Суточная потребность в витамине А для взрослого человека составляет 1, 5 мг (5000 ME).
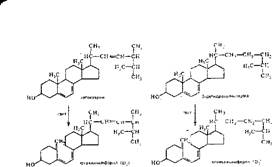
Химическая природа и биологически активные формы витамина А. Витамин А представляет собой непредельный одноатомный спирт, состоящий из р-иононового кольца и боковой цепи из двух остатков изопрена, имеющей первичную спиртовую группу. Витамин А имеет витамеры Аг и А2. Витамин А2. найденный у пресноводных рыб, имеет дополнительную двойную связь в р-ион'оновом кольце-: -
СН
HjC-^CH HjCvJ, I 1 Н3С< 1 I
НС
^—СН,
В организме ретинол (витамин А спирт) превращается в ретиналь (витамин А альдегид) и ретиноевую кислоту (витамин А кислота), т. е. происходит окисление спиртовой группы витамеров А! и А2 соответственно в альдегидную и карбоксильную. В тканях организма образуются производные витамина А — ретинилпальмитат и ретинилацетат (сложные эфиры ретинола с пальмитиновой и уксусной кислотами) и ретинилфосфат (фосфорный эфир ретинола). Витамин А и его производные находятся в организме в трд«с-конфигурации, за исключением сетчатки глаза, где образуются 11-цыс-ретинол и П-цис-ре- тнналь.
Известны три провитамина А — а-, р- н у-каротины, отличающиеся пе химическому строению и биологической активности. Наиболее активен р-каро- тин, который в слизистой кишечника подвергается окислению по центральной двойной связи с участием фермента каротиндиоксигеназы:
II, С Коротииаиокги/гнап С Н.
3\, сн, ларот. / 3
нгс—с 3! с—сн2
/ \ _Н _Н Н ____ н н * н_ н _н_ н / \
HjC\ /с—н—с—С_Н30-! - •' н " l-°~Н ~1 Н~ ^ / 2 н2с—с сн, < ^н3 сн3 С—CHj
При этом образуются две молекулы активного ретиналя. При распаде а- и у-каротннов, содержащих в отличие от p-каротина одно р-иононовое кольцо, образуется только по одной молекуле витамина А. Отсюда меньшая актив ность а- и у-каротинов по сравнению с р-каротином.
Биологической активностью обладают все формы витамина А: ретинол, ретиналь, ретиноевая кислота и их эфиропроизводные. •
Метаболизм. Для всасывания витамина А необходимы желчные кислоты, как и для всех липидов. В слизистой кишечника ретинол образует эфиры с жирными кислотами и транспортируется в составе хиломикронов. В плазме' ретинол связывается с ретннолсвязывающимся белком, находящимся во фрак ции с^-глобулинов, и доставляется в ткани. В сетчатке ретинол превращается в ретиналь, который входит в состав родопсина и играет важную роль в восприятии света. В печени эфиры ретинола депонируются. Часть ретинола в печени окисляется в ретиналь, а затем в ретнноевую кислоту, которая выводится с желчью в виде глюкуронидов.
Биохимические функции. Все формы витамина А (ретиналь, ретинол, ретиноевая кислота и их эфирные производные) регулируют следующие процессы:
1) нормальный рост и дифференнировку клеток развивающегося организма (эмбриона, молодого организма);
2) регуляцию деления и дифференцировкн быстро пролиферирующнх (делящихся) тканей — хряща и костной ткани, сперматогенного эпителия и плаценты, эпителия кожи и слизистых;
3) участие в фотохимическом акте зрения.
Ретиноевая кислота в акте зрения и функции размножения, т. е. нормальном развитии сперматозоидов в мужском организме и плаценты при беременности, не участвует. Она стимулирует рост костей н мягких тканей. Остальные формы витамина А обеспечивают все основные его биологические функции.
Окончательно механизм регуляции витамином А деления и дифференци-
|
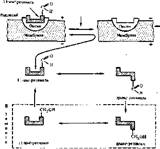
Рис 72 Схема зрительного акта ровки клеток яе выяснен. Возможно, это действие связано с запуском механизма репликации, а выраженное влияние на рост костей — с регуляцией синтеза хондроитинсульфата в клетках хряща. Многие метаболические функ ции витаминов А неясны. Детально изучено участие его в акте зрения. В этом процессе витамин А участвует в форме 11-цис-ретиналя, который входит в состав светочувствительных пигментов сетчатки глаза. У человека сетчатка имеет два типа клеток — палочки и колбочки. Палочки реагируют на слабое освещение (сумеречное, ночное зрение), а колбочки — на хорошее освещение (дневное зрение) и обеспечивают различение цветов (цветовое зрение). Палочки содержат зрительный пигмент родопсин, а колбочки — иодопсин. И тот, и другой являются сложными белками, состоящими из 11-^uc-ретиналя и белка опсина. Однако по строению белковой части родопсин и иодопсин различаются.
В зрительном акте можно выделить три процесса (рис. 72):
Ч) фотохимическая абсорбция света пигментом, который при этом изменяется;
2) образование нервного импульса в ответ на изменение пигмента;
3) регенерация исходного пигмента.
| г |
Кванты света, поглощаемые родопсином (или иодопсином), вызывают фотоизомеризацию 11-цис-ретиналя в гранс-ретиналь, после чего происходит диссоциация гранс-ретиналя и опсина и пигмент обесцвечивается. Поскольку пигменты встроены в мембраны светочувствительных клеток сетчатки, то фо- тонзомеризация ретиналя приводит к местной деполяризации мембраны и возникновению электрического импульса, распространяющегося по нервному волокну.
Регенерация исходного пигмента возможна прямым путем с участием рстинальизомеразы (это медленный процесс), протекающим на свету, или окольным путем в темноте. В темноте регенерация родопсина максимальная Она протекает через образование транс-ретинола, цис-ретинола и П-цис- ретинола. Последний вновь комплексируется с опсином. Отсутствие регенерации родопсина приводит к слепоте в ночное время или в сумерках.
Недостаточность витамина А. Наиболее ранним признаком недостаточности является нарушение темновой адаптации и ночная слепота. Кроме того, возможна задержка, роста в молодом возрасте, фолликулярный гиперкератоз (избыточное ороговение кожи, вызванное задержкой смены эпителия), сухость слизистых (гоже вследствие замедленного обновления эпителия), ксерофтальмия (сухость конъюнктивы глаза), помутнение роговицы и размягчение ее (кератомаляция), нарушение функции размножения (оплодотворяющей активности сперматозоидов).
Практическое применение. В медицинских целях используют природные препараты витамина А (содержащие смесь его биологических форм) и синтетические — ретинол ацетат и ретинолпальмитат. Они применяются для лечения гиповитаминозов как средство профилактики у людей, работа которых связана с напряжением зрения, для стимуляции роста и развития у детей, усиления регенерации плохо заживающих тканей, повышения сопротивляемости инфекциям, профилактики бесплодия.
Кальциферолы (витамин D)
Витамин D содержится в ряде продуктов животного происхождения: в печени, сливочном масле, молоке, а также в дрожжах и растительных маслах. Наиболее богата витамином D печень рыб. Из нее получают рыбий жир, используе-' мый для профилактики и лечения D-витаминной недостаточности.
Суточная потребность-в витамине D для детей колеблется от 12 до 25 мкг (500—1000 ME). Для взрослого человека нужны в десятки раз меньшие количества, чем для детей.
Химическая природа и биологически активные формы витамина D. Витамин D — групповое обозначение нескольких веществ, относящихся по химической природе к стеринам. Имеется несколько витамеров витамина D. Среди них наиболее активны эргокальциферол (D2), холекальциферол (D3) и дигид- роэргокальциферол (D4). Витамин D2 образуется из растительного предшественника (провитамина D) —эргостерина, а витамин D3— из 7-дегидрохоле- стерина (синтезирующегося в коже человека и животных) после облучения ультрафиолетовым светом растений или кожи человека и животных. Последний биологически более активен. Менее активные витамеры витамина D —D4, Ds, De, D7 образуются при облучении ультрафиолетом растительных предшественников (соответственно дчгидроэргостерина, 7-дегидроситосте- рина, 7-дегидростигмастерина и 7-дегидрокампестерина).
Однако ни эрго-, ни холекальциферолы биологически не активны и не могут выполнять свои регуляторные функции. Биологически активные формы их образуются в ходе метаболизма.
Метаболизм. Пищевые кальциферолы всасываются в тонком кишечнике с помощью желчных кислот. После всасывания они транспортируются в 'Со- 346
|
ставе хиломикронов в кровь и далее в печень. Сюда же с кроаью поступает и эндогенный холекальциферол. В-печени холекальциферол и эргокальцнферол подвергаются гидроксилированию в эндоплазм этическом ретикулуме с помощью 25-гидроксилазы холекальциферола. Образуются 25-гидроксихолекаль- циферол и 25-гидроксиэргокальциферол, которые принято считать основной транспортной формой витамина D. С кровью они переносятся в составе сле: циального кальциферол связывающего белка плазмы к почкам, где с участием i - гидроксил азы кальциферолов образуются 1, 25-дигидроксиксмьциферолы, которые и являются активной формой («гормональной») витамина D, регулирующей обмен кальция и фосфора в организме.
В тканях витамин D не накапливается, за исключением жировой ткани. Выводится он главным образом с калом в неизмененном или окисленном виде или в виде конъюгатов.
Биохимические функции. Биологическая активность 1, 25-гидрокснкальци- феролов в 10 раз превышает активность исходных кальциферолов. Витамин D регулирует транспорт ионов кальция и фосфора через клеточные мембраны и тем самым их уровень в крови. Эта регуляция основана, по крайней, мере, на трех процессах, в которых участвует витамин D:
1) транспорт ионов кальция и фосфата через эпителий слизистой тонкого кишечника при их всасывании;
2) мобилизация кальция из костной ткани;
3) реабсорбция кальция и фосфора в почечных канальцах.
Механизм действия на эти процессы витамина D предстааляется следующим
образом. Всасывание кальция в тонком кишечнике происходит путем облегченной диффузии с участием специального кальцийсвязывающего белка (СаСБ) и активного транспорта с помощью Саг+-АТФазы. 1, 25-Дигидрокси- кальциферолы индуцируют образование СаСБ и белковых компонентов Са2" 1" - АТФазы, возможно, действием на генетический аппарат клеток слизистой тонкого кишечника. Очевидно, подобная-стимуляция витамином D Са2+-АТФ- азы. мембран почечных канальцев приводит к реабсорбции в них ионов каль-
ция. Еще не ясны механизмы участия витамина D в трансмембранном переносе фосфата в кишечнике и почках и мобилизации кальции из костной ткани.
В целом действие витамина D выражается в повышении содержания ионов кальция и фосфатов в крови.
Недостаточность витамина D. Недостаточность витамина D проявляется в виде заболевания, названного рахитом. Развитие рахита у детей вызывается низким содержанием в пище, потребляемой детьми, витамина D, относительно меньшая, чем в старшем возрасте, возможность получать необходимую дозу ультрафиолетового облучения (для образования эндогенного витамина D) и меньшая чувствительность тканей, реагирующих на кальциферолы (очевидно, недостаток кальциферолсвязывающих рецепторов). При рахите заторможены все процессы, регулируемые витамином D, а именно: всасывание ионов кальция в фосфатов в кишечнике (хотя ребенок с молочной пищей получает их в достаточных количествах), реабсорбция их в почках. Вследствие этого уровень кальция и фосфора в крови снижается и нарушается минерализация костей, т. е. отложения минеральных веществ на вновь образовавшую колла- геновую матрицу растущих костей не происходит. Поэтому у страдающих рахитом наблюдается деформация костей скелета конечностей, черепа, грудной клетки.
Относительная недостаточность витамина D может быть и при нормальном его поступлении в организм. Она проявляется при заболеваниях печени н особенно почек, так как эти органы принимают участие в образовании, активных форм витамина D.
Гипервитаминоз D.< При приеме избыточных количеств витамина D у детей и взрослых развивается витаминная интоксикация. Она проявляется деминерализацией костей и их переломами. Уровень кальция и фосфатов в крови резко повышается (они извлекаются из костей, всасываются из кишечника и реабсорбируются в почках). Это приводит к кальцификации внутренних органов (из-за плохой растворимости кальция) — сосудов, легких, почек и др.
Практическое применение. В медицинской практике используются природные препараты витамина D (рыбий жир) и синтетические (эргокальцифе- рол нли холекальциферол).
В настоящее время налаживается производство активных метаболитов витамина D — 25-гидроксихолекальциферрла, обладающих большей антира- хитической активностью, которая к тому же не зависит от сопутствующих заболеваний печени.
Препараты витамина D применяются для профилактики и лечения рахита и для лечения других заболеваний (туберкулеза костей, кожи).
Нафтохиноны (витамин К)
Источником витамина К являются растительные (капуста, шпинат, отчасти корнеплоды и фрукты) и животные продукты (печень). Кроме того, он синтезируется бактериями тонкого кишечника. Суточная потребность в нем взрослого человека составляет примерно 2 мг.
Химическая природа и биологически активные формы витамина К. Витамин К по химической природе является хиноном с боковой изопреноидной цепью. Существует два ряда иафтохинонов, или витаминов К, — филлохиноны (витамины Кгряда) и менахиноны (витамины Кг-ряда): 348
о о
о < 1н1 I О I сн3 J,
Филлохиноны и их производные содержатся в растениях и поступают'с пищей, а менахиноны (сокращенно МК) образуются кишечными бактериями или в ходе метаболизма нафтохинонов в тканях организма. Синтетические препараты витамина К — менадион, викасол и синкавит являются производными 2-метил-1, 4-нафтохинона. В организме из них тоже образуются биологически активные менахиноны.
Метаболизм. Для всасывания пищевого витамина К в тонком кишечнике необходимы желчные кислоты и панкреатическая липаза. Водорастворимые препараты его в них не нуждаются. Транспорт всосавшегося витамина К происходит с хиломикронами. В плазме крови он связывается с альбуминами и накапливается в печени, селезенке и сердце. Большинство нафтохинонов пищевого и бактериального происхождения подвергается в тканях превращению с образованием менахинона, называемого МК-4 (который является витамином Кг-ряда). МК-4 служит, очевидно, основной биологически активной формой витамина К. Конечные продукты превращения витамина К выделяются с мочой.
Биохимические функции. Витамин К регулирует в организме процесс свертывания крови путем участия в образовании компонентов ее системы: фактора II (протромбина), фактора VII (проконвертина), фактора IX (фактора Кристмаса) и фактора X (фактора Стюарга). Витамин К участвует в превращении предшественника протромбина, называемого препротромбином, в протромбин. Этот процесс происходит в печени. Витамин К стимулирует у-кар- боксилирование остатков глутаминовой кислоты в молекуле протромбина, активируя микросомальную карбоксилазу. Образовавшийся протромбин связывается с фосфолипндами через ионы и подвергается ферментативному расщеплению с образованием тромбина. Последний автоматически зяпускает систему свертывания крови с образованием фибринового сгустка.
Недостаточность'витамина К- Признаками недостаточности витамйна К является повышенная кровоточивость, особенно при травмах. У взрослых людей кишечная флора полностью обеспечивает организм витамином К- У грудных детей (пока не развита кишечная флора) причиной гиповитаминоза может служить недостаток витамина К в пище. Основными причинами гиповитаминоза К являются: подавление кишечной флоры лекарственными средствами, заболевания печени н желчного пузыря, при которых нарушается образование желчных кислот (необходимых для.всасывания витаминов). К тому же печень служит местом образования активных форм витамина К, синтеза ряда факторов свертывания крови и превращения препротромбина в тромбин.
Практическое применение. В медицинской практике используются препараты витамина К[ и его синтетический аналог — викасол. Они применяются при кровоточивости или кровотечениях, связанных с понижением свертывания крови.
Токоферолы (витамин Е)
Источником токоферола для человека служат растительные масла: подсолнечное, кукурузное, хлопковое, оливковое. Особенно высоко его содержание в масле, полученном из зародышей пшеницы. Продукты животного происхождения, в том числе молочные, бедны токоферолом. Суточная потребность взрослого человека в токофероле примерно 20— 50 мг.
Химическая природа и биологически активные формы витамина Е. К витамину Е относятся метильные производные токола и токотриенола. Битами- ры Е обозначаются греческими буквами о-, 0у- и б-токоферолы и токотрие- нолы. По строению они очень близки. В их структуре имеется ароматический спирт токол и боковая изопреноидная цепь, которая у токоферолов полностью гидрирована, а у токотриенолов нет (рис. 73). Самым активным является а-токоферол, с которым обычно отождествляют Е-витаминную активность.
Метаболизм. Для всасывания пищевого токоферола необходимо присутствие, как и для всех жирорастворимых витаминов, липидов в качестве растворителей и желчных кислот как эмульгаторов. Всасывание их происходит в тонком кишечнике путем простой диффузии, затем в составе хиломикронов они транспортируются через лимфатические пути в кровь и с липопротеидами крови — в органы и ткани. В их клетках токоферол включается в состав мембран, где он и концентрируется. Наибольшее количество токоферола организ-

|
| Токоферол | Твкол | R' | R" | |
| о- | 5, 7, 8-траыетнл- | ац | сн3 | CHj |
| 5, 8-дамепт- | ац | И | ац | |
| ' 7- | 7> 8-вямет11Л | н | CHj | CHj |
| в- | 8-ыетнл- | н | Н | CHj |
| Токол | Н | н |
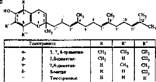 Рис. 73. Строение токоферолов (Л) и токотриенолов (б) по В. Б. Спкричеву
Рис. 73. Строение токоферолов (Л) и токотриенолов (б) по В. Б. Спкричеву
|
ма сосредоточивается в жировой ткани, печени и скелетных мышцах. Невсо- савшийся токоферол выводится с калом, а продукты метаболизма его в виде токофероновой кислоты и ее водорастворимых глюкуронидов — с мочой.
Биохимические функции. Гокоферол регульрусг интенсивность свободно- радикальных реакцчй в живых клетках, поскольку иреля'.хгвузт развитию цепных неуправляемых реакций пьрсксчдусго окисления ненасыщенных липидов в биологических мембранах.