Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Гликолиз — первый этап дыхания
|
|
Гликолиз — это универсальная последовательность реакций, общая для всех известных науке организмов, в ходе этих реакций глюкоза последовательно окисляется до пирувата. Окисление сопровождается образованием двух молекул НАДН и синтезом 2 молекул АТФ. Несомненно, что гликолиз — эволюционно самый ранний из известных способов получения энергии, и единственный для многих анаэробных бактерий. В аэробный метаболизм гликолиз включен как начальный этап окисления глюкозы.
Реакции гликолиза (рис. 4.3) протекают в цитозоле и катализируются ферментами, легко экстрагируемыми из клеток. Вначале богатая энергией, но стабильная молекула глюкозы фосфорилируется за счет АТФ в реакции (1), катализируемой гексокиназой. Образованный глюкозо-6-фосфат изомеризуется во фруктозо-6-фосфат (2) и еще раз фосфорилируется до фруктозо-1, 6-бисфосфата с использованием еще одной молекулы АТФ (3). Эта необратимая реакция, ключевая в регуляции гликолиза (см. подразд. 4.1.7), имеет у растений следующую особенность. Если у животных данную реакцию катализирует только один фермент —
АТФ-зависимая фосфофруктокиназа (3), то в клетках растений на этой стадии действуют сразу два фермента. Помимо АТФ-зависимой фосфофруктокиназы растения содержат также пирофосфатзависимую фосфофруктокиназу, которая катализирует фосфорилирование фруктозо-6-фосфата, используя не АТФ, а пирофосфат (4).
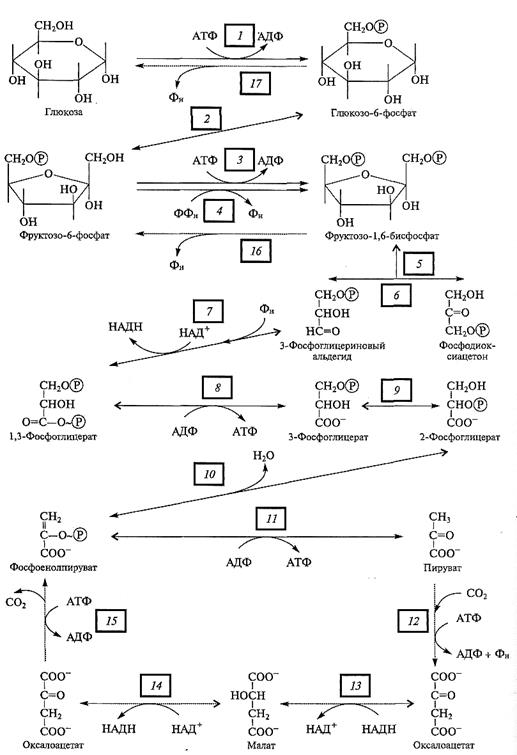
Рис. 4.3. Реакции гликолиза.
Реакции 1— 17 катализируют следующие ферменты: 1 — гексокиназа; 2 — фосфоглюкоизомераза; 3 — АТФ-зависимая фосфофруктокиназа (АТФ: В-фруктозо-6-фосфат-1-фосфотрансфераза); 4 — ФФн-зависимая фосфофруктокиназа; 5 — фруктозо-1, 6-бисфосфат альдолаза; 6 — триозофосфатизомераза; 7 — глицеральдегид-3-фосфат дегидрогеназа; 8 — фосфоглицераткиназа; 9 — фосфоглицеромутаза; 10 — енолаза; 11 — пируваткиназа (АТФ пируват-фосфотрансфераза); 12 — пируваткарбоксилаза; 13, 14 — НАД-зависимая малатдегидрогеназа; 15 — ФЕП-карбоксикиназа;
16 — фруктозо-1, 6-бисфосфатаза; 17 — глюкозо-6-фосфатаза; → — необратимая реакция;
↔ — обратимая реакция; ∙ ∙ ∙ ∙ ∙ ∙  — обходные реакции при обращении гликолиза;
— обходные реакции при обращении гликолиза;
фосфатная группа: 
Впервые этот фермент был обнаружен у пропионовокислых бактерий в начале 1980-х гг. в листьях ананаса (Ananas comosus), а затем и в других растительных объектах.
Цитозоль растительных клеток содержит пирофосфат (ФФН) в довольно высокой и устойчивой концентрации, который в ряде случаев выступает как энергетический эквивалент, или как донор фосфатных групп. ФФн-зависимая фосфофруктокиназа способна катализировать реакцию фосфорилирования фруктозо-6-фосфата в обоих направлениях, работая либо на синтез ФФН либо на его удаление. Предполагается, что таким образом контролируется пул ФФН в цитозоле.
Отрицательно заряженные фосфатные группы в молекуле фруктозо-1, 6-бис-фосфата облегчают ее распад при участии альдолазы на 3-фосфоглицериновый альдегид (3-ФГА) и фосфодиоксиацетон (ФДА) (5). Последний легко изомеризуется в ФГА (6). На этой реакции завершается первая стадия гликолиза: при использовании двух молекул АТФ глюкоза фосфорилируется и расщепляется на две половинки — фосфотриозы, которые в дальнейшем подвергаются одинаковым превращениям.
Вторая стадия гликолиза включает окислительно-восстановительные реакции, в которых образуются НАДН и АТФ. Окисление 3-фосфоглицеринового альдегида (7) катализирует глицералъдегид-3-фосфат дегидрогеназа. В ходе этойсложной реакции, которая идет через образование фермент-субстратного комплекса, восстанавливается НАДН и образуется
1, 3-дифосфоглицериновая кислота (1, 3-ФГК). Последняя представляет собой высокоэнергетическое соединение, в молекуле которого есть макроэргическая фосфатная связь. Синтез АТФ осуществляется при переносе фосфатной группы от 1, 3-ФГК на АДФ в реакции (8), катализируемой фосфоглицераткиназой. Таким образом, АТФ синтезируется в результате совместного действия двух реакций, в первой из которых макроэргическая связь образуется на молекуле окисляемого субстрата. Поэтому такой способ синтеза АТФ получил название субстратного фосфорилирования. В следующей реакции (9) 3-фосфоглицериновая кислота превращается в 2-фосфоглицериновую кислоту, от которой при участии енолазы отщепляется молекула воды (10). В результате этой реакции вновь образуется высокоэнергетическое соединение — фосфоенолпируват (ФЕП). Второе субстратное фосфорилирование связано с переносом фосфатной группы от ФЕП на АДФ, катализируемом пируваткиназой (11). Образованный пируват является конечным продуктом гликолиза.
У растений обнаружены ферменты, при участии которых возможен «обход» некоторых реакций гликолиза. Так, в клетках растений есть НАДФ-зависимая глицеральдегид-3-фосфат дегидрогеназа, катализирующая реакцию
3-ФГА + НАДФ+ + Н2О → 3-ФГК + НАДФН + 2Н+
В вакуолях некоторых растений (у Catharanthus roseus, Cupinus albus, Brassica nigra, Allium сера) обнаружена ФЕП-фосфатаза, катализирующая реакцию
ФЕП + Н2О  Пируват + Фн
Пируват + Фн
Оба альтернативных фермента активируются при фосфорном голодании.
Энергетический выход гликолиза небольшой. Если учесть, что на первом этапе активация глюкозы связана с расходом двух молекул АТФ, суммарный баланс можно выразить через уравнение
Глюкоза + 2НАД+ + 2ФН + 2АДФ→ 2Пируват + 2НАДН + 2Н+ + 2АТФ + 2Н2О
У растений гликолиз протекает не только в цитозоле, но и в пластидах. Пластидные и цитозольные изоформы кодируются разными ядерными генами и могут существенно различаться по своим свойствам. Пластидный гликолиз, связанный с мобилизацией крахмала, имеет место в гетеротрофных и фотосинтезирующих тканях. Однако в хлоропластах процесс, по-видимому, протекает только в темноте. Считается, что на свету активность пластидной изоформы
АТФ-зависимой фосфофруктокиназы подавлена из-за высокой концентрации АТФ (см. подразд. 4.1.7). Пирофосфатзависимая фосфофруктокиназа не обнаружена в хлоропластах и пластидах.
Предполагается, что образованные в гликолизе пируват, АТФ и НАДН используются в синтезе жирных кислот, который у растений протекает в пластидах. Следует также иметь в виду, что пластидный гликолиз может идти не до конца, так как такие соединения, как ФГА и ФГК, могут выходить из хлоропластов и включаться в гликолиз, протекающий в цитозоле.
При прорастании семян, если еще достаточно прочная наружная оболочка препятствует доступу кислорода, или при затоплении корней растений гликолиз завершается реакциями спиртового или молочнокислого брожения. При этом образованный ранее НАДН вновь окисляется при восстановлении пирувата соответственно до этанола или молочной кислоты. Обычно сначала при участии лактатдегидрогеназы образуется молочная кислота:
Пируват + НАДН + Н 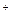
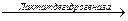 Лактат + НАД
Лактат + НАД
При накоплении молочная кислота подкисляет цитозоль, что необходимо для активации пируватдекарбоксилазы, которая при рН > 7, 0 находится в неактивной форме. При участии этого фермента на конечном этапе синтезируется этанол:
Пируват 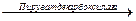 Ацетальдегид + СО2;
Ацетальдегид + СО2;
Ацетальдегид + НАДН + Н+  Этанол + НАД+
Этанол + НАД+
Этанол в отличие от молочной кислоты способен выходить из клеток в межклеточники, что менее опасно, чем накопление в цитозоле лактата. Брожение обеспечивает выживание растений ограниченное время в условиях недостаточного снабжения кислородом, т. е. при аноксии. По устойчивости к этому стрессу растения могут сильно различаться. Если некоторые болотные растения выживают в условиях аноксии в течение месяцев, то проростки ячменя или пшеницы не выдерживают и нескольких часов. Следует отметить, что в развивающихся пыльцевых зернах кукурузы (Zea mays) и табака (Nicotiana tabacum) спиртовое брожение имеет место в аэробных условиях и протекает наряду с дыханием.