Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Особенность растительных митохондрий — присутствие малик-энзима
|
|
Стабильная работы ЦТК во многом зависит от концентрации промежуточных соединений, а значит, и от процессов, связанных с их удалением или, наоборот, вовлечением в цикл. Многие интермедиаты цикла активно из него уходят и используются в различных реакциях в цитозоле (см. подразд. 4.1.8). В связи с этим важное значение имеют так называемые анаплеротические реакции, которые катализируют взаимопревращение продуктов гликолиза и ЦТК, и при необходимости «подкармливают» цикл, пополняя пулы промежуточных соединений.
Так, ФЕП-карбоксилаза катализирует синтез оксалоацетата из ФЕП:
ФЕП + НСО 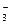
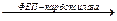 ОАА + Фн
ОАА + Фн
Как уже было показано (см. подразд. 4.1.4; рис. 4.3), оксалоацетат может быть также синтезирован из пирувата при участии пируваткарбоксилазы:
Пируват + С02 + АТФ 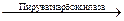 ОАА + АДФ + Фн
ОАА + АДФ + Фн
Малатдегидрогеназа катализирует легко обратимое взаимопревращение оксалоацетата и малата:
Малат + НАД+ 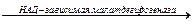 ОАА + НАДН + Н
ОАА + НАДН + Н 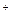
Особый интерес вызывает функционирование в растительной клетке малик-энзима, который катализирует окислительное декарбоксилирование малата:
Малат + НАД+ 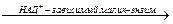 Пируват + СО2 + НАДН + Н
Пируват + СО2 + НАДН + Н
У растений малик-энзим присутствует не только в цитозоле, но и в митохондриях
(НАД+-зависимая изоформа) и хлоропластах (НАДФ+-зависимая изоформа). Наличие этого фермента в митохондриях делает возможным получать пируват из малата и тем самым обходить последнюю реакцию гликолиза (рис. 4.5).
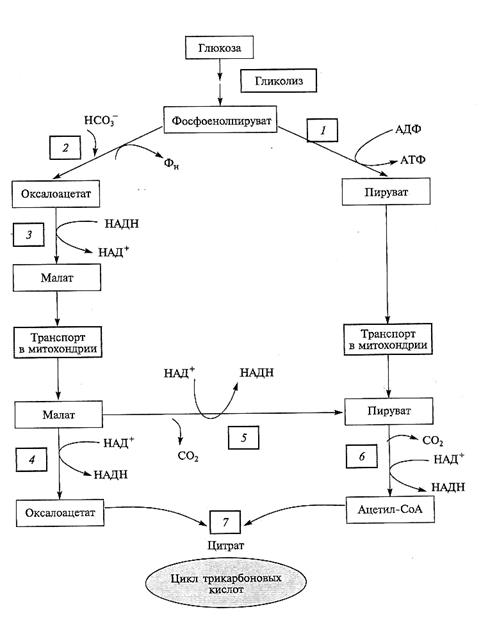
Рис. 4.5. Альтернативные пути образования пирувата в растительной клетке.
Образованный в гликолизе фосфоенолпируват (ФЕП) может метаболизировать с образованием пирувата или малата. ФЕП-карбоксилаза катализирует взаимодействие ФЕП с НСО с образованием оксалоацетата, который превращается в малат. Малат, как и пируват, поступает в митохондрии и может поддерживать ЦТК, превращаясь в пируват при участии малик-энзима. Реакции 1 — 7 катализируют ферменты: 1 — пируваткиназа; 2 — ФЕП-карбоксилаза; 3 — малатдегидрогеназа в цитозоле; 4 — малатдегидрогеназа в митохондриях; 5 — малик-энзим; 6 — пируватдегид-рогеназный комплекс; 7 — цитратсинтаза
Действительно, образованный в гликолизе ФЕП является субстратом сразу двух реакций. При участии пируваткиназы он превращается в пируват (1), а при участии ФЕП-карбоксилазы — в оксалоацетат (2). Последний легко воестанавливается до малата (5). Поступив в митохондрии, малат либо сразу включается в ЦТК, либо окисляется малик-энзимом до пирувата (5). На самом деле не совсем понятно, насколько этот альтернативный способ образования пирувата из малата используется in vivo в нормальной физиологической ситуации. Сообщалось, что этот путь активируется у некоторых растений при фосфорном голодании. Возможно, он включается при регуляторном подавлении активности пируваткиназы (см. подразд. 4.1.7). Тем не менее дублирование реакций, связанных с синтезом пирувата, отражает пластичность растительного метаболизма и, безусловно, расширяет их адаптивные возможности. Так, трансгенные растения табака (Nicotiana tabacum), лишенные пируваткиназы, выживали и поддерживали нормальный метаболизм за счет этого обходного пути.