Главная страница Случайная страница
КАТЕГОРИИ:
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Конверсия жиров в углеводы. Глиоксилатный цикл
|
|
В семенах некоторых растений жиры являются доминирующими запасными соединениями. К их числу относятся клещевина (Ricinus communis), арахис (Arachis hypogaea), рапс (Brassica napus), подсолнечник {Helianthus annuus), тыква (Cucurbita реро), соя (Glycine max), арабидопсис (Arabidopsis thaliana), пальма (Elaeis guineensis) и др.
В зависимости от вида растения жиры могут накапливаться в клетках семядолей (подсолнечник, рапс, арахис, соя) или в эндосперме (клещевина), т.е. в тканях, предназначенных «кормить» развивающийся проросток. При прорастании таких семян действует особый путь метаболизма, через который жиры превращаются в сахарозу, и она транспортируется в органы развивающегося проростка, где используется как субстрат дыхания и как источник углеводов для биосинтезов (например, клеточной стенки).
На пути превращения жиров в углеводы в тканях эндосперма или семядолей действует глиоксилатный цикл, ферменты которого локализованы в особых органеллах — глиоксисомах.
Глиоксисомы принадлежат к пестрому семейству микротелец, которое у растений представлено глиоксисомами и пероксисомами. По мере развития проростка глиоксисомы либо исчезают вместе с эндоспермом, либо превращаются в типичные пероксисомы при зеленении семядолей.
Уникальная конверсия жиров в углеводы включает три этапа: окисление жирных кислот, глиоксилатный цикл и глюконеогенез. Все три этапа связаны между собой через взаимодействие четырех компартментов — жировых телец, глиоксисом, митохондрий и цитозоля (рис. 4.10, 4.11). Обычно растения запасают триацилглицеролы, которые в клетке находятся в виде жировых телец, или олеосом. Олеосомы отделены от водной фазы мембраной, состоящей из одного слоя фосфолипидов. Такая необычная структура возникает при образовании олеосом: жиры синтезируются в полостях ЭПР, откладываются между двумя монослоями мембраны, а затем отщепляются в виде капель. Специальные белки, олеозины, покрывают их поверхность и предотвращают слипание. При прорастании семян жиры подвергаются гидролизу при участии липазы с образованием жирных кислот и глицерола. Глицерол может превращаться в триозофосфат и участвовать в гликолизе, а свободные жирные кислоты поступают в глиоксисомы, которые как правило находятся с олеосомами в тесном контакте. Здесь начинается процесс
β -окисления, в результате которого от жирной кислоты последовательно отщепляются 2С-фрагменты с образованием ацетил-СоА. Процесс сопровождается восстановлением НАД+ и образованием перекиси. Образованный НАДН может выноситься из глиоксисом через малат-оксалоацетат-челнок, а перекись разлагается каталазой (см. подразд. 4.3.2).
Ацетил-СоА является субстратом глиоксилатного цикла, который представляет собой модифицированный ЦТК (рис. 4.10).
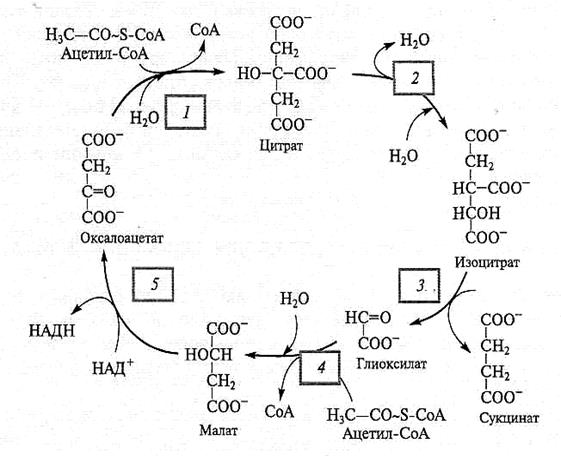
Рис. 4.10. Глиоксилатный цикл.
Реакции 1— 5 катализирут ферменты: 1 — цитратсинтаза; 2 — аконитаза; 3 — изоцитратлиаза; 4 — малатсинтаза; 5 — НАД-зависимая малатдегидрогеназа. Все ферменты, участвующие в цикле, локализованы в глиоксисоме, за исключением, возможно, аконитазы. Есть данные, что аконитаза в прорастающих семенях клещевины и тыквы является цитозольным ферментом
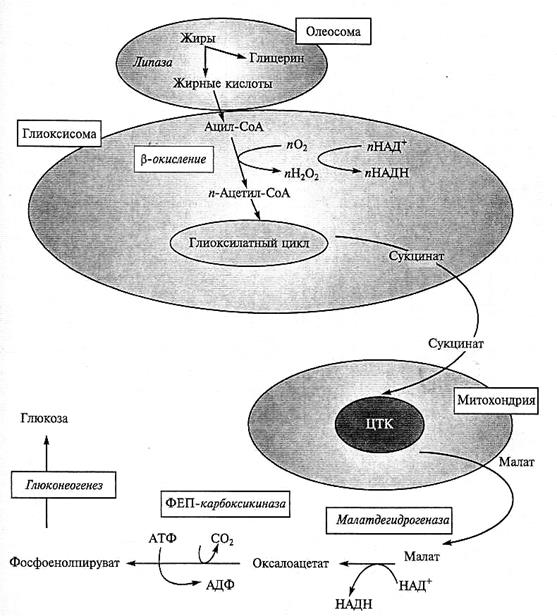
Рис. 4.11. Взаимодействие компартментов в процессе превращения жиров в углеводы при прорастании семян масличных растений.
Запасенные в виде жировых капель жиры подвергаются гидролизу липазой. Свободные жирные кислоты конденсируются с коэнзимом А и образуют ацил-СоА. Последний поступает в глиокси-сомы, где происходит процесс
β -окисления, в котором от жирной кислоты последовательно отщепляются ацетильные остатки в виде ацетил-СоА. Ацетил-СоА метаболизирует через глиоксилатный цикл, продуктом которого является сукцинат. Сукцинат поступает в митохондрии, где под действием ферментов ЦТК превращается в малат. Образованный в цикле малат выходит из митохондрий и метаболизирует в цитозоле с образованием фосфоенолпирувата. Фосфоенолпируват через глюконеогенез превращается в 6С-углеводы
Три фермента — цитратсинтаза, аконитаза и малатдегидрогеназа, общие для глиоксилатного цикла и ЦТК, представлены в глиоксисомах другими изоформами. Две первые реакции глиоксилатного цикла идентичны двум первым реакциям ЦТК. Ацетил-СоА конденсируется с оксалоацетатом с образованием цитрата, который затем превращается в изоцитрат при участии аконитазы. Последующие две реакции специфичны для глиоксилатного цикла и катализируются двумя ферментами, уникальными для растений. В первой из них изоцитратлиаза катализирует расщепление изоцитрата с образованием сукцината и глиоксилата. Сукцинат уходит из цикла, а глиоксилат в реакции, катализируемой малатсинтазой, вновь конденсируется с еще одной молекулой ацетил-СоА с образованием малата. Замыкает цикл реакция окисления малата до оксалоацетата, идентичная аналогичной замыкающей реакции ЦТК. Таким образом, в результате пяти реакций из двух молекул ацетил-СоА (ацетильных остатков) синтезируется сукцинат, который уходит из глиоксисом и используется в синтезе углеводов. Согласно простейшей схеме сукцинат поступает в митохондрии, где включается в реакции ЦТК и восстанавливается до малата. Малат выносится из митохондрий в цитозоль и окисляется до оксалоацетата при участии малатдегидрогеназы. ФЕП-карбоксикиназа катализирует реакцию, в которой оксалоацетат превращается в ФЕП. Фосфоенолпируват, ключевой метаболит гликолиза, может быть превращен в глюкозу и сахарозу в процессе глюконеогенеза (см. подразд. 4.1.4).
Глиоксилатный цикл функционирует не только в прорастающих семенах масличных растений, но и в стареющих листьях, а также при созревании пыльцы. Предполагается, что гены, кодирующие ферменты глиоксилатного цикла, имеются у всех растений, но экспрессируются не во всех тканях и не на всех стадиях онтогенеза. При старении листьев пероксисомы, вероятно, трансформируются в глиоксисомы, и в них начинает функционировать глиоксилатный цикл, с помощью которого метаболизирует ацетил-СоА, образованный при деградации липидов мембран. Образующиеся сахара в дальнейшем уходят из стареющих листьев в другие органы.